
恐竜
| 恐竜類 Dinosauria | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||||||||||||||||||||||||||||||||
| 地質時代 | |||||||||||||||||||||||||||||||||||||||||||||
| 約23,140万- 現代 (中生代後期三畳紀 - 現代) | |||||||||||||||||||||||||||||||||||||||||||||
| 分類 | |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| 学名 | |||||||||||||||||||||||||||||||||||||||||||||
| Dinosauria Owen, 1842 | |||||||||||||||||||||||||||||||||||||||||||||
| 和名 | |||||||||||||||||||||||||||||||||||||||||||||
| 恐竜類[1] | |||||||||||||||||||||||||||||||||||||||||||||
| 英名 | |||||||||||||||||||||||||||||||||||||||||||||
| Dinosaur | |||||||||||||||||||||||||||||||||||||||||||||
| 目 | |||||||||||||||||||||||||||||||||||||||||||||
恐竜(きょうりゅう、英: Dinosaur)は、恐竜類(きょうりゅうるい[1]、学名: Dinosauria)に属する爬虫類の一群である。爬虫綱での分類階級を示す場合は恐竜上目とされる[2]。全長30メートル程ある巨大なものから、数十センチメートル程しかない小型なものまで、様々な属が生息していた[3]。
恐竜の進化に関する正確な起源と時期は不明であるが、2億4330万年から2億3323万年前の三畳紀中期に出現したものが起源とされている。恐竜は2億130万年前に発生した三畳紀〜ジュラ紀間における大量絶滅を生き延び、その後のジュラ紀と白亜紀を通して陸生脊椎動物の頂点に立ったが、白亜紀~新生代古第三紀間における大量絶滅により、鳥類を除くすべての種が絶滅した。なお一部の非鳥類型恐竜がK-Pg境界を乗り越え、続く暁新世の地層から産出した事例も報告されている[4]。ただし異論も多く、2023年現在有力な説にはなっていない(詳細は暁新世の恐竜を参照)。
概要
[編集]最初の恐竜の化石は19世紀初頭に発見され、 1841年にリチャード・オーウェンによってこれらの「偉大な化石トカゲ」を指すために「恐竜 (dinosaur) 」(「恐ろしいトカゲ」を意味)という呼称が造られた。それ以来、化石恐竜の骨格標本は世界中の博物館の主要な目玉であり、恐竜は大衆文化の永続的な一部になっている。現存する生物に比べ、非常に大きな体格と怪物的な外観を有する種が多いため、『ジュラシック・パーク』に代表されるSF作品に登場することも多く、またゴジラをはじめとする多数の創作モンスターのモチーフにも用いられた。
恐竜という言葉は、分類学的・形態学的・生態学的な観点から、非常に細かい「分類」である。現代まで生存している唯一の生物種である鳥類は10,700種以上が確認されており、脊椎動物の中で最も多様なグループの1つとなっている。食性はさまざまで、植物食性・肉食性の双方の種が確認されている。
恐竜は現存する種(鳥類)と化石によって、すべての大陸に分布していたことが分かっている。20世紀の前半まで、鳥類が恐竜として認識される前は、恐竜は鈍重な変温動物であるという認識が一般的だった。しかし、1970年代におこった恐竜ルネッサンスにより、一部の恐竜は代謝が高く、社会的相互作用に多くの適応がある活発な動物であることが示されている。また、現状で確認されている種は鳥類・非鳥類ともに全て卵生であり、大半の種に巣を作る習性があったことが判明している。現代の鳥類は、後期ジュラ紀に獣脚類の中から派生した羽毛恐竜を起源とし、約6600万年前の白亜紀と古第三紀の間の大量絶滅を生き延びた唯一の恐竜系統であることが化石記録から判明している。そのため、恐竜は「鳥類型恐竜」と、鳥類以外の恐竜である絶滅した「非鳥類型恐竜」に分けられる。
恐竜はすべてのグループにおいて角やとさかなどの外見構造が発達しており、一部の絶滅したグループは骨格を発達させたものが確認されている。唯一の現存群である鳥類系統は、大半の種が飛行を行うために小柄であるが、恐竜(非類および鳥類)の大半は鳥類に比べ体が大きく、中でも最大の竜脚類であったアルゼンチノサウルスは長さ39.7メートル、高さ18メートルに達したと推定されており、史上最大の陸上動物であった。化石を証拠として、古生物学者は900以上の異なる属と1,000以上の異なる種の非鳥類型恐竜を特定した。非鳥類恐竜が一様に巨大であるという誤解は、骨が大きく頑丈であるほど化石になるまで保存されやすいという保存バイアスに基づいている。しかし、多くの恐竜は非常に小さく、長さが約50センチメートルのものも存在していた。
名称
[編集]
英語: dinosaur はギリシャ語: δεινός, deinos 「恐ろしい」 + σαῦρος, sauros 「とかげ」[注 1]の合成である。1842年、それまでに発見されていた3種の化石爬虫類(イグアノドン・メガロサウルス・ヒラエオサウルス)の新しい分類名と一括し、リチャード・オーウェンによって「恐竜」という命名を名付けされた[5]。
オーウェンはδεινός の語を英語でfearfully great[6][7]、つまり恐ろしいほど大きな[8]という意味で用いたという。語根 -saur- には「竜」をあてるのが通例である。「○○サウルス」といえば「恐竜の名前」として認識されがちだが、実際にはトカゲ型の動物や爬虫類には、現生のものであっても「-saurus」と学名の付いたものが少なくない。例えばエリマキトカゲの学名はChlamydosaurus kingiiであり、カタカナに直せば"クラミドサウルス・キンギイ"となる。
定義
[編集]通俗的には「恐竜」という言葉は往々にして「大昔の爬虫類」という程度の把握しやすいイメージで認識されており、同じ地質時代に生息していた各種の爬虫類のほかに、古生代に生息していた一部の非哺乳類型の単弓類なども含めた概念として呼ばれる場合が少なくない。「恐竜」とは系統的に異なり、大衆的に恐竜の一群としてイメージされやすい翼竜・魚竜・首長竜・モササウルスなどは一切含まない独立した分類群である。恐竜展や子供向けの恐竜図鑑などではこれらの古代生物たちまで含めて展示・掲載するものがよく見られるため、初心者にとっては混同し易い。
古典的定義
[編集]恐竜という分類群はそのもっとも際立った特徴をして「直立歩行に適した骨格をもった爬虫類」と呼ぶことができ、ほぼすべて地上棲である[9]。翼竜は恐竜や鳥類・ワニと同じく主竜類に属し、恐竜とは「姉妹群」の関係にあたる。
古典的分類によれば、恐竜は爬虫綱 - 双弓亜綱 - 主竜形下綱に属し、分類階級は上目とされてきた。なお、系統樹に基づく分岐学的観点から単に「恐竜」と呼んだ場合、学術的には「鳥類」を含める(後述)。このため、上記の分類群(恐竜から鳥類を除いたグループ)を指す上では、より厳密な「非鳥類型恐竜(non-avian dinosaur)」の使用が、学術論文を中心に見られる[10][11]。ただし一般に「恐竜」と言えば鳥類を除いたものを指すケースが多く、依然分類群としても簡便で有用である[11]。よって本項では特に言及のない限り、「恐竜」と言えば「非鳥類型恐竜」を指すものとする。
分岐学的定義
[編集]分岐学の観点から、「現生鳥類とトリケラトプス(Triceratops)を含むグループの最も近い共通祖先より分岐したすべての子孫」が定義として頻繁に用いられる[12][10]。
この意味は実際にクラドグラムを見ると分かりやすい。
以下は、Nesbitt (2011)に基づく主竜類のクラドグラムの例:
| 主竜類 |
| ||||||||||||||||||||||||||||||||||||||||||

上の系統図で、現生鳥類は竜盤類の獣脚類に含まれ、トリケラトプスは鳥盤類の一属である。要するに、前述の定義の意図するところは概して「竜盤類と鳥盤類、それぞれの動物の共通祖先から分岐したすべてのもの」[11]であり、「現生鳥類」「トリケラトプス」は、それぞれ竜盤類、鳥盤類における代表例として任意に挙げられたにすぎない[注 2]。
よって同様のグループを、例えば「恐竜 (dinosauria)」の命名のきっかけとなった2属を挙げ、「メガロサウルス(Megalosaurus)とイグアノドン(Iguanodon)を含むグループの、最も近い共通祖先より分岐したすべての子孫」[13]と表すこともできる。
また、より厳密に3つの系統を用いて「トリケラトプス (Triceratops horridus)、サルタサウルス(Saltasaurus loricatus)、イエスズメ(Passer domesticus)の、最も近い共通祖先より分岐したすべての子孫」(つまり、恐竜=鳥盤類+獣脚類+竜脚形類)[14]とする意見もある。
これらの定義では必然的に、獣脚類の一群である鳥類を恐竜(より詳細には、竜盤類の中の獣脚類、コエルロサウリアに属すマニラプトラに含まれる)に含めることになる。このため、鳥を除いた恐竜を表すために、「非鳥類型恐竜 (non-avian dinosaur)」の用語が使用される。また特に、鳥を除いた獣脚類を表す語として、「非鳥類型獣脚類 (non-avian theropod)」も頻繁に用いられる。
なお、恐竜類の中のクレードについては上記のクラドグラムと異なりオルニトスケリダやフィトディノサウルス類を設ける説もある[15]。
以下は、オルニトスケリダのクラドグラムの例。
| 恐竜形類 |
| ||||||||||||||||||||||||||||||
以下は、フィトディノサウルス類のクラドグラムの例。
| 恐竜類 |
| ||||||||||||||||||

特徴
[編集]中生代三畳紀に現れ、中生代を通じて繁栄した。多様な形態と習性のものに適応放散し、陸上動物としては非常に大きくなったものもあったが、約6,600万年前の白亜紀と新生代との境に多くが絶滅した。なお、アラモサウルスなどの一部の属については、この後もしばらく生き延びていた可能性を主張する研究者もいる[16]。以前より鳥類は恐竜(の一部)から進化したものだという見方があった[17]が、獣脚類の一部は現在も鳥類として繁栄しているとする説が主流となっている[18]。
特徴的な派生形質
[編集]
恐竜は単系統群と考えられており[10][11]、その系統を特徴づける派生形質は非常に多い。以下に主要なものを列挙する[11]。
(主竜類、さらに鳥頸類や恐竜型類の形質をもった上で)
- 後前頭骨(postfrontal)を二次的に失う。
- 上腕骨の三角筋稜(delto-pectoral crest)が発達する。
- 腸骨、恥骨、坐骨で構成された骨盤に、大腿骨がはまり込む場所である寛骨臼(かんこつきゅう)が貫通している。
- 仙椎を構成する骨が3個以上(ワニやトカゲなど、多くの爬虫類ではより少ないことが多い)。
- 脛骨前面に距骨突起が成立する。
など。
これらのいくつかは、特に二足歩行に適応した結果として生じた形質と考えられ、恐竜が、一般的に想像されるトカゲのような「爬虫類」とは異なる運動機能を持っていたことを示している。かつては、二足歩行は前肢を武器とするために進化したと考えられていたが、これには疑問点もある[19]。
進化史
[編集]登場と初期進化
[編集]恐竜は、祖先的な双弓類から進化した群で、直接的祖先は初期主竜類(かつては槽歯目としてまとめられていた)中の一群、鳥頸類の一群とされる[20]。このグループには恐竜の他には翼竜などが含まれている。このグループは、初期段階から二足歩行へと移行しつつある形態を持っており、最初期の恐竜は既に二足歩行を獲得していた。なお恐竜は三畳紀当初は生態系の10%前後を占める程度の弱小勢力であったが、三畳紀後期から緩やかに獣弓類やクルロタルシ類から主役の座を譲り受け、以降の繁栄を築いた[21]。
非鳥類型恐竜の絶滅
[編集]非鳥類型恐竜は白亜紀末期に絶滅した。恐竜はよく関心を持たれる動物群であり、ことさらその絶滅の原因に関する仮説は多い。しかし、ある系統の「絶滅」とは、生物の進化において普遍的なプロセスであり、中生代を通じていくつもの恐竜の系統が絶滅してきたことにも留意する必要がある[22]。
K-Pg境界(以前はK-T境界と呼ばれた)の大量絶滅は、恐竜のみならず数多くの動植物を巻きこんだという意味で大規模な絶滅であり、事実、K-Pg境界における恐竜の絶滅に関する科学的な研究は長い間なされてこなかった[11]。
絶滅の主要因に関する仮説には以下などのものがある。
最も有力とされているのは巨大隕石の衝突である。1980年、地質学者のウォルター・アルバレスとその父で物理学者のルイス・アルバレスは、世界的に分布が見られる中生界白亜系と新生界古第三系を境する粘土層(通称K-T境界層)に含まれるイリジウムの濃度が他の地層の数十倍であり、かつ、イリジウムは地殻にはほとんど存在しないことから、これが隕石の衝突によってもたらされたものであると考え、大量絶滅の原因を隕石の衝突に求めた[23]。その後、1991年メキシコ・ユカタン半島に、直径180キロメートルの巨大クレーター(チチュルブ・クレーター)が再発見され、このクレーターを形成した隕石の衝突が恐竜絶滅の原因だとする説が提唱された[24]。この説では、地球規模の大火災で生態系が破壊され、衝突後に生じた塵埃が大気中に舞い、日光を遮断することで起きた急速な寒冷化が絶滅の原因であると主張された(ただし異論を唱える学者もいる[25])。2010年、Peter Schulte他40名の研究者も、チチュルブ・クレーターを形成した衝突が大量絶滅を引き起こしたと結論づけた[26]。
しかし一方で、衝突で大気中に浮遊した微小粉塵量を過大評価しているとし、寒冷化よりもむしろ衝突で大気中に浮遊した粉塵・衝突による巨大な森林火災の煤煙などが地表への太陽光をさえぎった結果、地上や海中の生態系が破壊され、食物連鎖の底辺の光合成を行う生物の様相が大きく変わり、隕石衝突の直接の影響を生き抜いた恐竜たちも餌の不足により絶滅したとする説明が提示されている。なお、東北大学は前述の説を支持する研究結果を出している[27]。また、隕石が南側に数百キロずれて衝突していたら恐竜は今も生き残っていたかもしれないとする研究結果もある[28]。
隕石説と反対に、イリジウムの起源を地球内部に求め、当時活動していたデカントラップなどの火山活動が大量絶滅の原因であるとする「火山説」も複数の研究者により唱えられている。また、両者を組み合わせ、隕石衝突が5万年に渡る連続的な火山活動を引き起こし、それが絶滅につながったという「連動説」も存在する[29]。
過去には伝染病説、裸子植物から被子植物への植物相の変化(草食恐竜の食物が無くなった)、原始的な哺乳類による恐竜の卵乱獲説など諸説もあったが、現在ではかえりみられない。これら諸説は、恐竜のみの絶滅の原因を考察したものであり、白亜紀末期の恐竜を含めた数多くの動植物の絶滅の原因の説明になっていないからである。
当初の衝突による「衝突の冬」(寒冷化)が原因では、なぜ同時期に存在した両生類や爬虫類などが絶滅を免れたかという疑問が残ったが、現在でも二酸化炭素による濃度上昇に伴う気温上昇、塵による太陽光の遮断、硫酸エアロゾルによる太陽光遮断と酸性雨などについては確証がなくよくわからないとする意見も強い。
鳥類
[編集]鳥類が生き延びた恐竜の一種であることが学術的に認められるようになったことで、「恐竜は絶滅していない」「絶滅を逃れた恐竜が鳥類である」といった観点を立てることができる[30]。
生物学的特徴
[編集]
現代に生きる鳥の系統を除いても、恐竜は長期間にわたって陸上で繁栄した一群であり、その形態は多様であった。
その身体のサイズも、ニワトリほどの大きさのものから、陸上においては最大級のものまでさまざまであった。最大のものは竜脚類で、その中でも判明している範囲ではスーパーサウルスが最も大きい。これら竜脚類は、水棲のクジラ類を除けば地球の歴史上最も大きな動物であり、陸棲動物では地球史上最大である[31]。マラアプニサウルスやブルハトカヨサウルスのようにさらに巨大なサイズに達したと推測される竜脚類も記載されたものの、化石が現存しないため不確かな状態にある[32][33]。現存するもので部分的な標本としてはバロサウルスもしくはスーパーサウルスに属するとされる大型標本があり、一部の推定では全長40 - 50 メートルの範囲に達したと推測された[34][35]。
体重についてはブラキオサウルス科のギラファティタンなどが80トン以上だったとする説もある。ただし、この数値はやや過大であるという見方も存在する[36]。
恐竜の祖先である初期主竜類は肉食性であり、エオラプトル、コエロフィシスなど最初期の恐竜も肉食の捕食者であった[37]。しかし、原竜脚下目など比較的初期段階から草食へと移行しつつあるグループも出現している[38]。
姿勢・歩行
[編集]恐竜はそれ以外の多くの爬虫類とは異なり、胴体の直下に四肢を持つ。この特徴は、側方型の四肢に比べて体重を支えるのに都合がよく、大型化したグループが出現する素地となったとする考えがある。また、歩行の際に身体を捻る必要がないため、軽快な移動を可能にしている。
この特徴はやや遅れて哺乳類も獲得しているが、異なる点としては、恐竜は二足歩行の種が多い点である。これは、二足歩行が初期主竜類から受け継いだ、祖先的な形態だからである。初期主竜類の中でもユーパルケリアなどは身体の作りが軽快であり、一時的な二足歩行を可能としていた(現生のエリマキトカゲに近い動きだった)。竜脚類や鳥盤類の一部の様な四足歩行の恐竜は、体重の増加等の理由で二次的に四足歩行に復帰したものである。このためか、四足歩行の恐竜でも体重の大半は後足が支える形となっている[39]。この特徴的な2足歩行かつ爪先立ちという姿勢は、その後の恐竜の繁栄にとって欠かせない要因の一つとなった[40]。
元より軽量な小型恐竜は別として、跳躍走行(脚が全て地面から離れ、一瞬身体が宙に浮く移動方法)が可能だった大型恐竜はアロサウルスのような大型獣脚類のみだったとされている[41]。
恐竜の二足歩行はヒトとは異なり、後足を中心に長い尾によって上半身と下半身のバランスをシーソーのようにとっていたと考えられている。恐竜の巨大な尾はバランスを取るための必然であり、ティラノサウルスおよび近縁の属に見られる縮小した前肢は、巨大化した頭部と釣り合いを取るためだとされる[要出典]。恐竜に関する研究が未熟だった1970年代以前では、二足歩行、四足歩行ともに恐竜は尾を引きずりながら歩くと考えられていたが、恐竜ルネッサンスのあった1970年代以降では間違いとされ、現在の体格が提唱された[要出典]。よってその前後では、児童書や図鑑などに掲載されている恐竜の絵柄が大きく変わっている。
一方、胴体の横から足が生えている側方型の爬虫類、例えばトカゲは胴体をくねらせて歩行する、いわば爬行を行っている。この方法では四肢を側方に突き出した姿勢で身体を持ち上げているため、エネルギー効率が悪い。また肺を圧迫するために呼吸が阻害され、長時間の走行を困難にしている[42]。
カメは四肢が側方に生えておりながらも胴体が甲羅に覆われて可動性を持たないため、爬行を行えない。そのため四肢により胴体を持ち上げて歩行を行っているが、歩行能力は一般に優れているとは言えず、歩行速度の遅い動物の代表格扱いされている。またワニは、トカゲに比べて胴体の可動性がやや乏しいために[43]、胴体をくねらさない。また、短距離ではあるとはいえ、ギャロップで走行することが可能なものも存在する[44]。ただし、ワニは恐竜に近縁なグループであり、祖先は胴体直下に直立した四肢を持っていた。そのため、かれらの四肢を突き出した姿勢は半水生の生態に適応した二次的なものである。またカメもワニなどに近縁な主竜形類であるため、祖先は胴体直下に四肢を持っていたが、甲羅の発達にともない二次的に四肢が側方へと突き出す形となったとする仮説もある[45]。
恐竜の二足歩行形態は、現在では子孫の鳥類へと受け継がれている。しかし、祖先と異なる点としては、尾が短縮したことで重心が前方へと移ったため、大腿骨がほぼ身体に対して水平に保持されていることである[46]。そのため、歩行は膝関節を中心としたものとなっている[47]。
肌・色
[編集]
恐竜がどのような色をしていたのかは化石からは不明で、科学的根拠がないため明らかではなく、図鑑などに載っている恐竜の色は制作者のセンスで決められて、爬虫類と同様の茶色やくすんだ緑色など地味なものが多かった[48]。その後、鳥類との関係が認知され、羽毛をもつ恐竜が発見されるに従い、カラフルな恐竜の復元画も登場してきている。
2008年、ヤコブ・バンターらは恐竜の羽の化石中に含まれるメラニン色素を解析することにより、オリジナルの色がどのようなものであったか判別することに成功した[49][信頼性要検証]。今後、羽に覆われた保存状態のよい皮膚組織の化石が発見されれば、恐竜がどのような色をしていたのか解明することが可能と主張している。
羽毛
[編集]
そして1990年代以降、中国の白亜紀の地層で羽毛をもった、現在の鳥類と羽毛のない恐竜の間を埋める、羽毛のある恐竜の化石が相次いで発見され、系統関係が明らかになってきた。羽毛をもった恐竜には、シノサウロプテリクス・プロターケオプテリクス・カウディプテリクス・ミクロラプトル・ディロングなどがある。[要出典]
羽毛をもった恐竜のグループの存在から空を飛ぶ鳥類と恐竜の進化の関係が明確になった。このように現在では、「鳥類は恐竜の獣脚類の一種である」という説がほぼ定説となった[52]。
気嚢式呼吸
[編集]
前気嚢 肺 後気嚢
古生代ペルム紀末(P-T境界)の大量絶滅により、陸上で多様化や大型化を遂げていた単弓類の一群など、数多くのグループが絶滅した。こうして空白地帯となった陸上の生態系に、さまざまなグループが競って進出し、ニッチを埋めていったと思われる。その中の一つが恐竜の祖先であった。他の競合者となったのが、鳥頸類の姉妹群でワニの祖先を含むクルロタルシ類及び、前時代からの残存勢力で哺乳類の祖先を含む単弓類であった。
ペルム紀初期の大気中の酸素濃度は35%に達したとされ、横隔膜による呼吸を採用した哺乳類型爬虫類は高酸素下では特に支障はなかった[53]。出現当初の恐竜は比較的小型であり、ほぼ同時期に現れた哺乳類共々、陸上の脊椎動物相においてそれほどの割合を占めていなかったとされる。しかし、三畳紀末期には、恐竜の多様性が増大する。この理由のひとつとして呼吸器系の進化が提唱されている。すなわち進行する乾燥と低酸素化の環境の中、哺乳類は横隔膜を使った肺呼吸をより発達させて効率的な酸素交換を実現し、一方の恐竜は、それより更に高効率の呼吸システムを獲得していく。それが、鳥類にも見られる気嚢である。この気嚢を獲得したことで、恐竜は高い酸素摂取能力を獲得することになる。また、この気嚢が骨の中に入り込むことで中空の含気骨となり、骨格自体の軽量化にも貢献することとなった。
酸素濃度35%のペルム紀以降は、リグニンの分解能を獲得した菌類による木材の分解により酸素濃度は低下しジュラ紀後期の2億年前には酸素濃度は12%まで低下した。気嚢は、横隔膜方式よりも効率的に酸素を摂取できる機能があり、低酸素下でもその機能を維持し繁栄することができた。競合する哺乳類型爬虫類は低酸素下でその種の大部分が絶滅することとなった[53]。
恒温性
[編集]初めて恐竜が見つかったときには、爬虫類であるとして変温動物と考えられていた。それに異を唱え、「恐竜は恒温動物である」とした研究者にはジョン・オストロムやその弟子のロバート・T・バッカーなどがいる。彼らを含む研究者の一部は、恐竜を含む主竜類、特に小型の獣脚類は温血動物であったと主張している。[要出典]
しかしながら脳の発達の程度、骨に年輪が見られることなどから恒温性を否定する説も存在した。しかし、骨の年輪は草食哺乳類にも存在することが判明しており、この点からも恒温性を否定する根拠とはなり得ない[54][55]。
またエドウィン・ハリス・コルバートらがアメリカアリゲーターの総排泄口に温度計を差し込み、体温を計測する実験を行った結果、大型の個体程体温の変化が緩やかであるという結論が得られた[56]。故に、「大型の竜脚類などでは容積が大きいので結果的に体温を体内に保つことが出来る「慣性恒温性」で体温を保っていた」とする主張もあり、研究が続けられている。
生態・社会性
[編集]
非鳥類型恐竜は絶滅しており、その行動を直接見ることは出来ないため、生態は不明瞭な点が多い。数少ない物証としては、ヴェロキラプトルとプロトケラトプスが戦っている状態で見つかった化石や、鳥類のように丸まって眠っている姿勢で発見されたメイ・ロンの化石、同種の歯型が多数残り共食いをしていたことが推定されるマジュンガサウルスの化石がある[要出典]。
恐竜の行動の多くは足跡や巣の状態から類推することができ、化石のみで情報が乏しいながらも、骨格から推測される筋肉、足跡の計測などから、おおよその歩行速度を求める試みも一部にはある[要出典]。
また、子供を育てる、群れを作って共同で生活をするなど、現在見られる哺乳類動物と類似する社会性をもった恐竜もいたと考えられている。社会性をもつと、捕食動物にもよるが、捕食者が近づいて来た場合の警告がしやすい。しかし、これらはまだ研究者の間で議論中の論点であり、異論も少なくない。
繁殖~孵化
[編集]恐竜も他の動物と同じく交尾をし、産卵する。求愛行動を行った可能性がある。 また、一度に多くの卵を産むことができるため、繁殖効率も高くなる。
- 獣脚類
主に獣脚類は浅く掘った穴に卵を産み付け、土砂や植物を被せるといった単純な巣や、卵が、巣の中央に空間ができるようにドーナツ状に、2~3層に重ねて産み付け、すべての卵を産み終えると土砂や植物で覆おおい、卵のない巣の中央に親がうずくまって卵を保護していたと考えられているもの、巣が、地面を浅く掘ったU字型で、ふちが低く盛り上がっており、卵は2つ1組で、柔らかくした巣の中の地面に、卵のとがった方から突き刺すようにして産みつけられたものがある。また、産卵後には、現代の鳥類のように卵を抱いて温めていたと考えられている[57]。
- 竜脚類(主にティタノサウルス形類)
竜脚類は主に、後ろ脚を使って一方向に穴を掘り、浅く細長い巣を作って卵を産み落としていたとす推測されている。一回の産卵で産む卵の数が100個を超える場合もあったと考えられている[57]。
- 鳥脚類
鳥脚類の中でもマイアサウラは、すりばち状の巣に最大で40個の卵を産みつけ、植物を卵の上にかぶせていたと考えられている。巣の中の卵の配置には、規則性はなくバラバラだった[57]。
地中に巣を掘り、地中に卵を産む方法は、卵を保護し孵化まで温度や湿度を調節することができる利点があった。また、一度に多くの卵を産むことができるため、繁殖効率も高くなった[58]。
研究史
[編集]発見
[編集]
学術的な記録としては1677年、イギリス、オックスフォード大学のアシュモリアン博物館にてロバート・プロット(Robert Plot)による大腿骨の膝関節部分の記載が最初と思われる。これはオックスフォード州中期ジュラ紀の地層より発掘されたもので、おそらくメガロサウルスのものと推測される(詳細なスケッチを残し、標本は現存していない)が、この時代に恐竜は知られておらず、プロット自身はゾウのような大型の動物の骨と考えていた[59][60]。

1815年頃にはウィリアム・バックランドが新たな化石を入手している。バックランドはこの化石がどのような動物に属するのか判断がつかなかったようだが、1818年にはジョルジュ・キュヴィエがオックスフォードを訪れ、この化石が大型の爬虫類のものであることを指摘している。これに基づきバックランドは1824年にはTransactions of the Geological Society誌上にて論文を発表し、断片的な下顎、いくつかの脊椎骨や腸骨、後肢の一部の化石を記載。Megalosaurus(メガロサウルス)と命名した[61]。
1822年にはロンドン地質協会でギデオン・マンテルが、ある"植物食性と思われる動物の歯の化石"について発表を行った。これは当初、同協会に所属していたバックランドや、既に比較解剖学の大家として知られていたキュヴィエらから、サイの歯かあるいは魚のものであるとの評価を得た。しかし後年の精査により、彼らもこれが大型の爬虫類のものであると認め、1825年、マンテルはこの歯の化石をもとにIguanodon(イグアノドン)を記載した[62]。
1842年には、リチャード・オーウェンにより、メガロサウルス、イグアノドン、ヒラエオサウルスを内包するグループとして"Dinosauria"(恐竜)の名称が初めて用いられた[5]。
化石戦争
[編集]鳥との関係
[編集]
1861年、ドイツのゾルンホーフェンにおける後期ジュラ紀の地層より発掘された化石に基づき、Archaeopteryx(始祖鳥)が初めて報告された[63]。始祖鳥は、それまで鳥に特有とされていた羽毛を持ちながら、発達した歯や手指、長い尾を持つなど、爬虫類のような特徴を多く保持していた。1868年には、トマス・ハクスリーがこの化石に基づき、鳥の祖先が恐竜であったと述べている[64]。1926年に出版されたThe Origin of Birdsでゲルハルト・ハイルマン(Gerhard Heilmann、1859–1946)は、鳥は恐竜ではなく、恐竜よりも原始的な主竜類である槽歯類から分岐したと主張した[65]。ハイルマンの主張の根拠は、当時恐竜には鎖骨がないと考えられていたことである。ドロの不可逆則により、恐竜で消失した鎖骨が鳥類で復活して叉骨に進化することはなく、恐竜と鳥の類似点はいずれも収斂進化によるとしたのである。実は、この本の出版より前に鎖骨のある恐竜の化石がいくつか発掘されていたのだが、これらの化石での鎖骨の存在は当初見落とされていた[注 3]。ハイルマンは徹底的な調査を行っていたため、この本はこの分野での古典となり、この考え方はその後の40年間、ほぼすべての進化生物学者に受け入れられていた[67]。1936年に鎖骨のある原始的な獣脚類セギサウルスが発掘されたが、その後も恐竜には鎖骨がないとするのが正当な考え方であった[68]。1969年にジョン・オストロムらがデイノニクスを記載し(次項で詳述)、ふたたび鳥の祖先として恐竜が挙げられるようになった[69]。その後もジャック・ゴーティエによる分岐学的手法の発達[70][71]や、新たな祖先的鳥類やマニラプトラ類をはじめとする化石の発見が相次ぎ、現在では(本記事で幾度も述べられた通り)恐竜と鳥との関係はほとんど疑いのないものとなっている[52]。
恐竜ルネッサンス
[編集]1969年に記載された小型の獣脚類であるDeinonychus(デイノニクス)に基づく一連の研究は、それまでの「大型でのろまな変温動物」という恐竜のイメージを、恒温性で活動的な動物へと大きく覆した。1970年代に続いたオストロムやその学生であるロバート・バッカーらの主導による一連のパラダイムシフトはdinosaur renaissance(恐竜ルネッサンス)と呼ばれた。これ以降、恐竜の行動や生態、進化や系統に関する多種多様な研究が増えていった。
2000年代以降の主な報告
[編集]指の謎
[編集]恐竜と鳥との関係は確実視されてはいたが、両者の間では前肢の指に齟齬があった。恐竜では鳥の系統に近づくにつれ、五本指のうち第5,4指が退縮する(つまり第1,2,3指が残る)傾向があるのに対し、現在の鳥の指は位置関係上、第2,3,4指であることが発生学的に観察されるからである[72]。この問題についてギュンター・ワグナー(Günter Wagner)らは、非鳥類型恐竜から鳥への進化上の段階で、前肢の指の位置関係のシフトが起こったと説明した[73][74]。また田村らはニワトリの胚を用い、発生の過程で指の原基となる細胞が、位置関係のシフトを起こすことを示した[75]。
これに対して2009年には第1指、第5指の退化的な(つまり2,3,4指の発達した)リムサウルス(Limusaurus)が報告される[76]など、いまだ指には謎がある。
軟組織の発見
[編集]2000年から2003年、アメリカ・モンタナ州の約6800万年前の地層で見つかった恐竜化石から、ティラノサウルス・レックスの化石化していない軟組織が発見された。ただし2009年頃には、それが現代のバクテリアの粘液に由来するものではないかという疑惑が浮上している[要出典]。
他にもカモノハシ竜のミイラ化石とされる「ダコタ」など、軟組織が含まれているのではないかと考えられる化石は存在する。
別種か、それとも?
[編集]トリケラトプスとトロサウルス[77][78]、パキケファロサウルスとスティギモロクとドラコレックスのように、これまで別属と考えられていた恐竜が、成長段階や雌雄の差なのではないかとする研究がある。いずれも反論があり、共通見解は得られていない。
復元
[編集]恐竜の習性は化石では直接確認できないことが多いが、足あとの化石や生息地が習性を予想する手がかりになる。マイアサウラのように子育てを継続的に行っていた証拠が確認された例などもあるが、恐竜の行動にはなお不明な点が多く、現在恐竜展などで展示されている恐竜の姿や行動は、鳥類の習性からの予想や似た生態的地位にある現生大型動物からの類推による部分も大きい。
かつて恐竜は、ワニのような皮膚をもっていたと考えられており、実際に鱗が保存された化石も発見されている。その後鳥類との類縁関係が注目されるようになり、一部の種においては羽毛をもった化石も発見されたところから、中にはある種の鳥類のような色鮮やかな羽毛をもつものがいた可能性も考えられている。ただし、図鑑等で見られる恐竜の皮膚や羽毛の色模様等は全て現生爬虫類または鳥類から想像されたもので、実際の皮膚がどんな色だったかはほとんど不明である。皮膚自体が残った、ミイラ状態の化石は発掘されているが、質感はともかく色や模様は化石として残らないからである。
下位分類
[編集]恐竜の定義としては、寛骨(いわゆる骨盤)に大腿骨がはまり込む場所である寛骨臼(かんこつきゅう)が貫通していること、仙椎を構成する骨が3個以上である、などである。下位の分類としては、骨盤の形状により、竜盤類 Saurischiaと鳥盤類 Ornithischiaに大別される[79]。竜盤類は恥骨が前方へと伸びる、トカゲなど他の爬虫類に似た骨盤を持っていた。対して鳥盤類は鳥類に似た、恥骨が腸骨にそって後方へと伸びた形状であった。両者の分岐は約2億2900万年前に起きたと見られている[80]。
恐竜の分類はジャック・ゴーティエが1986年に分岐分析法により作成した系統樹により大きく変貌している。鳥類が恐竜(獣脚類)から分岐したこともこのときに示されている。
分類学的には竜盤類と鳥盤類を恐竜とする。翼竜、魚竜、首長竜など恐竜と同時代にはさまざまな大型爬虫類が出現したが、分類上、これらは恐竜には含まれない。この3グループの系統上の位置は爬虫類を参照。現生の動物では鳥類が最も近い生物であり、ワニ類がそれに次ぐ。カメ類はこれより遠く、ヘビ・トカゲ類(有鱗目)とはさらに遠い系統関係になる。
なお、新種の恐竜の化石が発見される、同種だと思われていた恐竜が別種だった、逆に別種だと思われていたが同種だった、骨格から体の特徴が改められるなどの新発見が相次いでおり、毎年のように最も有力な学説は変化している。即ち恐竜研究は現在も速い速度で進展しつづけているのであり、現在最も有力な知見が将来も有力であり続ける保証は存在しない。たとえばブロントサウルスは、1980年代頃まではほぼ必ず恐竜図鑑に登場する代表的な恐竜とされていたが、これは学名のBrontosaurusをカタカナ表記したものであり、現在はApatosaurusに学名が変更されたためカタカナ表記もアパトサウルスへ変化した。




上位分類
[編集]竜盤類 Saurischia
[編集][要出典]
- ?エオラプトル科 Eoraptoridae - エオラプトル
- ?ヘレラサウルス類 Herrerasauria
- ヘレラサウルス科 Herrerasauridae - ヘレラサウルス、レヴェルトラプトル
- スタウリコサウルス科 Staurikosauridae - スタウリコサウルス (三畳紀、南米)
- グアイバサウルス科 Guaibasauridae - グアイバサウルス
- 新獣脚類 Neotheropoda
- ケラトサウルス下目 Ceratosauria
- コエロフィシス上科 Coelophysoidea
- ゴジラサウルス Gojirasaurus
- コエロフィシス科 Coelophysidae
- プロコンプソグナトゥス亜科 Procompsognathinae - プロコンプソグナトゥス
- コエロフィシス亜科 Coelophysinae - コエロフィシス、サルトプス、メガフォサウルス
- ?ハルティコサウルス科 - ハルティコサウルス、ディロフォサウルス
- ネオケラトサウルス類(新ケラトサウルス類) Neoceratosauria
- コエロフィシス上科 Coelophysoidea
- テタヌラ下目 Tetanurae
- ?クリオロフォサウルス Cryolophosaurus
- メガロサウルス上科 Megalosauroidea
- アヴェテロポーダ類 Avetheropoda(新テタヌラ類 Neotetanurae)
- カルノサウルス類 Carnosauria
- アロサウルス上科 Allosauroidea
- モノロフォサウルス Monolophosaurus
- シンラプトル科 Sinraptoridae - シンラプトル、ヤンチュアノサウルス、スゼチュアノサウルス
- アロサウルス科 Allosauridae - アロサウルス、クリオロフォサウルス、サウロファガナクス、アクロカントサウルス
- カルカロドントサウルス科 Carcharodontosauridae - カルカロドントサウルス、ギガノトサウルス、マプサウルス

Allosaurus fragilis(アロサウルス・フラギリス)の爪のレプリカ
- アロサウルス上科 Allosauroidea
- コエルロサウルス類 Coelurosauria
- ?オルニトレステス科 Ornitholestidae -オルニトレステス
- コンプソグナトゥス科 Compsognathidae - コンプソグナトゥス、シノサウロプテリクス(中華竜鳥)
- コエルルス科 Coeluridae - コエルルス、オルニトレステス
- ティラノサウルス上科 Tyrannosauroidea-エオティラヌス、
- オルニトミモサウルス類 Ornithomimosauria
- オルニトミムス科 Ornithomimidae - オルニトミムス、ストルティオミムス、ガリミムス

マニラプトル類の一種、ヴェロキラプトルの復元。
- オルニトミムス科 Ornithomimidae - オルニトミムス、ストルティオミムス、ガリミムス
- マニラプトル類 Maniraptora
- アルヴァレスサウルス科 Alvarezsauridae - モノニクス、アルヴァレスサウルス、パタゴニクス
- オヴィラプトロサウルス形類 Oviraptoriformes
- オヴィラプトロサウルス類 Oviraptorosauria
- アビミムス科 Avimimidae - アビミムス
- カウディプテリクス科 Caudipteridae - カウディプテリクス
- カエナグナトゥス上科 Caenagnathoidea
- カエナグナトゥス科 Caenagnathidae - カエナグナタシア、キロステノテス、エルミサウルス
- オヴィラプトル科 Oviraptoridae - オヴィラプトル、アジャンキンゲニア、インキシヴォサウルス
- テリジノサウルス類 Therizinosauria
- テリジノサウルス上科 Therizinosauroidea
- オヴィラプトロサウルス類 Oviraptorosauria
- 原鳥類 Paraves
- ペドペンナ Pedopenna
- ?スカンソリオプテリクス科 Scansoriopterygidae - スカンソリオプテリクス、エピデクシプテリクス
- デイノニコサウルス類(デイノニクス竜類)Deinonychosauria
- 鳥類 Aves(現生鳥類の先祖)
- カルノサウルス類 Carnosauria
- ケラトサウルス下目 Ceratosauria


- テコドントサウルス科 Thecodontosauridae - テコドントサウルス
- 原竜脚下目 Prosauropoda
- ?アンキサウルス科 Anchisauridae - アンキサウルス
- プラテオサウルス科 Plateosauridae - プラテオサウルス、ウナイサウルス
- メラノロサウルス科 Melanorosauridae - メラノロサウルス
- マッソスポンディルス科 Massospondylidae - マッソスポンディルス、ルーフェンゴサウルス
- 竜脚下目 Sauropoda
- ?ヴルカノドン科 Vulcanodontidae - ヴルカノドン
- 真竜脚亜目(エウサウロポーダ類) Eusauropoda
- ?イービノサウルス
- マメンチサウルス類
- マメンチサウルス科 Mamenchisauridae - マメンチサウルス、チュアンジェサウルス
- ケティオサウルス類
- ケティオサウルス科 Cetiosauridae - ケティオサウルス、ロエトサウルス
- ジョバリア科 - ジョバリア
- オメイサウルス科 Omeisauridae - オメイサウルス
- 新竜脚類 Neosauropoda
- ハプロカントサウルス Haplocanthosaurus
- ディプロドクス類 Diplodocimorpha
- レッバキサウルス科 Rebbachisauridae
- ディプロドクス上科 Diplodocoidea
- Flagellicaudata
- ディプロドクス科 Diplodocidae
- ディクラエオサウルス科 Dicraeosauridae - ディクラエオサウルス、アマルガサウルス、レッバキサウルス
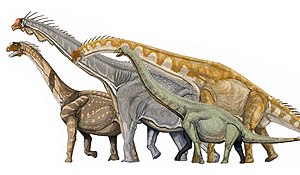
代表的なマクロナリア類の復元。左からカマラサウルス、ブラキオサウルス、ギラファティタン、エウヘロプス
- Flagellicaudata
- マクロナリア類 Macronaria
- カマラサウルス類 Camarasauromorpha
- カマラサウルス科 Camarasauridae - カマラサウルス
- ティタノサウルス形類 Titanosauriformes
- ?アンデサウルス科 - アンデサウルス、マラウィサウルス
- ブラキオサウルス科 Brachiosauridae - ブラキオサウルス
- ティタノサウルス上科 Titanosauria
- カマラサウルス類 Camarasauromorpha

鳥盤類 Ornithischia
[編集]ファブロサウルス科 Fabrosauridae - ファブロサウルス、レソトサウルス、テクノサウルス
ゲナサウリス類(頬竜類) Genasauria - エキノドン
- 装楯亜目 Thyreophora
- スクテロサウルス科 - スクテロサウルス
- ユーリ脚類(寛脚類) Eurypoda
- スケリドサウルス科 Scelidosauridae - スケリドサウルス
- ステゴサウルス類(剣竜下目) Stegosauria
- 曲竜下目(鎧竜下目) Ankylosauria
- ステゴペルタ族 Stegopeltini - グリプトドントペルタ、ステゴペルタ
- ノドサウルス科 Nodosauridae - ノドサウルス、サウロペルタ、エドモントニア、ドラコペルタ
- ポラカントゥス科 Polacanthidae - ポラカントゥス、ガーゴイロサウルス、ガストニア
- アンキロサウルス科 Ankylosauridae - アンキロサウルス、エウオプロケファルス、サイカニア
- 角脚類 Ceraopoda(新鳥盤類 Neornithischia)
- ヘテロドントサウルス科 Heterodontosauridae - ヘテロドントサウルス
- 鳥脚亜目 Ornithopoda
- ゼフィロサウルス科 Zephyrosauridae - ゼフィロサウルス
- オスニエリア科 Othnieliidae - オスニエリア
- 新鳥脚類 Neornithopoda(正鳥脚下目 Euornithopoda)
- ヒプシロフォドン科 Hypsilophodontidae - ヒプシロフォドン
- イグアノドン類 Iguanodontia
- ムッタブラサウルス Muttaburrasaurus
- 真イグアノドン類 Euiguanodontia
- Rhabdodontidae - ラブドドン
- ドリオサウルス科 Dryosauridae - ドリオサウルス、ヴァルドサウルス
- アンキロポレクシア Ankylopollexia
- カンプトサウルス科 Camptosauridae - カンプトサウルス、ドラコニクス
- イグアノドン上科 Iguanodontoidea
- イグアノドン科 Iguanodontidae - イグアノドン、オウラノサウルス、フクイサウルス
- プロバクトロサウルス科 Probactrosauridae - プロバクトロサウルス
- ハドロサウルス科 Hadrosauridae - ハドロサウルス、アナトティタン、コリトサウルス、ランベオサウルス、パラサウロロフス、マイアサウラ、シャントゥンゴサウルス
- 周飾頭亜目 Marginocephalia
- ステノペリクス Stenopelix
- ヘテロドントサウルス科 Heterodontosauridae - ヘテロドントサウルス、ゲラノサウルス、ラナサウルス
- 堅頭竜下目 Pachycephalosauria
- ホマロケファレ上科 Homalocephaloidea
- パキケファロサウルス科 Pachycephalosauridae - パキケファロサウルス、ステゴケラス
- ホマロケファレ科 Homarocephalidae - ホマロケファレ
- ホマロケファレ上科 Homalocephaloidea
- 角竜下目 Ceratopia
- プシッタコサウルス科 Psittacosauridae - プシッタコサウルス
- 新角竜類 Neoceratopsia
主な恐竜の分類群の時間範囲
[編集]ホルツによる主な恐竜の分類群の時間範囲の表。

日本で発掘された主な化石
[編集]- 北海道芦別町…ティラノサウルス類
- 北海道小平町…ハドロサウルス類の後ろ足
- 北海道夕張市…ノドサウルス類の頭骨の一部
- 北海道むかわ町…ハドロサウルス科、カムイサウルス・ジャポニクス(むかわ竜)[86]
- 岩手県岩泉町…竜脚類(通称モシリュウ)
- 福島県南相馬市…足跡(ジュラ紀)
- 福島県広野町…鳥脚類(通称ヒロノリュウ)、ティラノサウルス類(通称フタバリュウ)[87]
- 福島県いわき市…竜脚類(通称ヒサノハマリュウ)、鳥脚類
- 群馬県神流町(旧中里村)…スピノサウルス類の歯、オルニトムス類(通称サンチュウリュウ)、獣脚類、足跡
- 長野県小谷村…足跡(ジュラ紀)
- 富山県富山市(旧大山町)…イグアノドン類の歯、獣脚類、足跡
- 岐阜県白川町…竜脚類、足跡
- 岐阜県高山市(旧荘川村)…イグアノドン類、ヒプシロフォドン類の歯、鳥脚類、獣脚類
- 石川県白山市(旧白峰村)…竜脚類(通称ハクサンリュウ)、獣脚類(通称オピラプトロサウルス類)、イグアノドン類(通称シマリュウ)、アルバロフォサウルス・ヤマグチオロウム(角脚類)[88]、ティラノサウルス類(通称カガリュウ)[89]、足跡
- 福井県勝山市…フクイサウルス・テトリエンシス(イグアノドン類)[90]、フクイラプトル・キタダニエンシス(メガラプトル類)[91]、フクイティタン・ニッポネンシス(竜脚類)[92]、コシサウルス・カツヤマ(イグアノドン類)[93]、フクイベナートル・パラドクサス(コエルロサウルス類)、フクイプテリクス・プリマ (鳥群)、ティラノミムス・フクイエンシス (オルニトミモサウルス類)[94]、獣脚類、鳥脚類、足跡
- 福井県大野市(旧和泉村)…獣脚類の歯、足跡
- 三重県鳥羽市…ティタノサウルス類(通称トバリュウ)、足跡
- 兵庫県丹波市.丹波篠山市…タンバティタニス・アミキティアエ(ティタノサウルス形類)、ヒプノヴェナトル・マツバラエトオオエオルム(コエルロサウルス類)、ササヤマグノームス・サエグサイ(新角竜類)、角竜類、ティラノサウルス類、鎧竜類など
- 兵庫県洲本市…ヤマトサウルス・イザナギイ(ハドロサウルス類)ランベオサウルス類
- 徳島県勝浦町…イグアノドン類の歯
- 山口県下関市吉母…足跡
- 福岡県北九州市…獣脚類
- 福岡県宮若市(旧宮田町)…獣脚類(ワキノサウルス・サトウイ)
- 長崎県長崎市…獣脚類[95]
- 熊本県御船町…ティラノサウルス類、オルニトミムス類、テリジノサウルス類、ドロマエオサウルス類、アンキロサウルス類、ハドロサウルス類、ミフネリュウ、足跡
- 熊本県天草市…獣脚類、イグアノドン類、鳥脚類、竜脚類、足跡
- 熊本県八代市…獣脚類(アロサウルス形類)、肋骨[96]
- 鹿児島県薩摩川内市(旧下甑村、旧鹿島村)…獣脚類の歯、イグアノドン類の歯、角竜類の歯
博物館
[編集]脚注
[編集]注釈
[編集]出典
[編集]- ^ a b 冨田幸光, 對比地孝亘, 三枝春生, 池上直樹, 平山廉, 仲谷英夫「恐竜類の分岐分類におけるクレード名の和訳について」『化石』第108巻、日本古生物学会、2020年、23-35頁、doi:10.14825/kaseki.108.0_23。
- ^ 巌佐庸・倉谷滋・斎藤成也・塚谷裕一編「生物分類表」『岩波 生物学辞典 第5版』岩波書店、2013年、1531–1666頁。
- ^ 小林快次『恐竜の学校』小林快次監修、ニュートンプレス、2023年、20頁。ISBN 978-4-315-52725-4。
- ^ Fassett, J. E.; Zielinski, R. A.; Budahn, J. R. (2002). “Dinosaurs that did not die: evidence for Paleocene dinosaurs in the Ojo Alamo sandstone, San Juan Basin, New Mexico.”. Geological Society of America Special Papers 356: 307-336. doi:10.1130/0-8137-2356-6.307.
- ^ a b Owen, R. (1842). "Report on British Fossil Reptiles." Part II." Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841.London: John Murray. pp.: 60-204
- ^ ASIN 0253333490, The Complete Dinosaur ISBN 978-0253333490
- ^ “恐竜は「恐ろしい竜」ではない”. 恐竜の楽園 (2011年1月19日). 2012年12月10日時点のオリジナルよりアーカイブ。2017年7月2日閲覧。
- ^ “悩ましい翻訳語―科学用語の由来と誤訳”. はてなブログ. はてな (2011年6月14日). 2017年7月2日閲覧。
- ^ 小畠郁生(監修)、西川寛、高岡昌江、三品隆司『恐竜の大常識』ポプラ社、2003年、14頁。ISBN 4591078353。
- ^ a b c Dinosauria2nd 2004, pp. [, 要ページ番号],
- ^ a b c d e f Fastovsky & Weishampel 2006, pp. [, 要ページ番号],
- ^ Benton MJ: Origin and relationships of Dinosauria. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. Zweite Auflage. University of California Press, Berkeley 2004, S. 7-19, ISBN 0-520-24209-2.
- ^ Olshevsky, G. (2000). "An annotated checklist of dinosaur species by continent". Mesozoic Meanderings 3: 1–157.
- ^ Sereno, P. C. (2005). “The logical basis of phylogenetic taxonomy”. Syst. Biol. 54 (4): 595–619. doi:10.1080/106351591007453. PMID 16109704.
- ^ ダレン・ナイシュ、ポール・バレット 著、小林快次、久保田克博、千葉謙太郎、田中康平 監訳 吉田三知世 訳『恐竜の教科書 最新研究で読み解く進化の謎』創元社、2019年2月20日、45頁。ISBN 978-4-422-43028-7。
- ^ 読売新聞、2011年2月5日22時8分配信。
- ^ “鳥は恐竜から進化した ー 論争についに終止符 | Nature ダイジェスト | Nature Portfolio”. www.natureasia.com. 2022年1月24日閲覧。
- ^ “驚きの恐竜展を開催、もはや鳥展、米NYで”. natgeo.nikkeibp.co.jp. 2022年1月24日閲覧。
- ^ The functional origin of dinosaur bipedalism: cumulative evidence from bipedally inclined reptiles and disinclined mammals(W Scott Persons:2017)
- ^ ヴォード 2008, p. 253
- ^ A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs(Randall B Irmis:2007)
- ^ 「濱田隆『恐竜学への誘い』」『恐竜学がわかる。』朝日新聞社〈AERA MOOK〉、2001年、[要ページ番号]頁。ISBN 4022741163。
- ^ Alvarez, L.W.; Alvarez,W., et al. (1980). “Extraterrestrial Cause for the Cretaceous-Tertiary Extinction”. Science 208 (4448): 1095-1108. doi:10.1126/science.208.4448.1095.
- ^ ジェームズ・ローレンス・パウエル『白亜紀に夜がくる-恐竜の絶滅と現代地質学』寺嶋英志、瀬戸口烈司(訳)、青土社、2001年、[要ページ番号]頁。ISBN 4791759079。
- ^ ムー (雑誌)2008年1月号、学研[出典無効]
- ^ Schulte, Peter (2010). Laia Alegret, Ignacio Arenillas, Jose A. Arz, Penny J. Barton, Paul R. Bown, Timothy J. Bralower, Gail L. Christeson, hilippe Claeys, Charles S. Cockell, Gareth S. Collins, Alexander Deutsch, Tamara J. Goldin, Kazuhisa Goto12, Jose M. Grajales-Nishimura, Richard A. F. Grieve, Sean P. S. Gulick, Kirk R. Johnson, Wolfgang Kiessling, Christian Koeberl, David A. Kring, Kenneth G. MacLeod, Takafumi Matsui, Jay Melosh, Alessandro Montanari, Joanna V. Morgan, Clive R. Neal, Douglas J. Nichols, Richard D. Norris, Elisabetta Pierazzo, Greg Ravizza, Mario Rebolledo-Vieyra, Wolf Uwe Reimold, Eric Robin, Tobias Salge, Robert P. Speijer, Arthur R. Sweet, Jaime Urrutia-Fucugauchi, Vivi Vajda, Michael T. Whalen, Pi S. Willumsen. “The Chicxulub Asteroid Impact and Mass Extinction at the Cretaceous-Paleogene Boundary”. Science 327 (5970): 1214-1218. doi:10.1126/science.1177265.
- ^ 恐竜やアンモナイト等の絶滅は「小惑星衝突により発生したすすによる気候変動」が原因だった東北大学公式ホームページ
- ^ 小惑星が少しずれていたら…恐竜生存? 東北大など発表朝日新聞
- ^ “Asteroid that killed dinosaurs also intensified volcanic eruptions - study” (英語). the Guardian (2015年10月2日). 2020年12月26日閲覧。
- ^ 鳥は「恐竜の子孫」ではありません、恐竜なんです | ギズモード・ジャパン
- ^ コルバート, モラレス & ミンコフ 2004, pp. 199–200
- ^ Carpenter, Kenneth (2018-10-19). “Maraapunisaurus fragillimus, N.G. (formerly Amphicoelias fragillimus), a basal Rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado” (英語). Geology of the Intermountain West 5: 227–244. doi:10.31711/giw.v5.pp227-244. ISSN 2380-7601.
- ^ Paul, Gregory S.; Larramendi, Asier (2023-04-11). “Body mass estimate of Bruhathkayosaurus and other fragmentary sauropod remains suggest the largest land animals were about as big as the greatest whales” (英語). Lethaia 56 (2): 1–11. doi:10.18261/let.56.2.5. ISSN 0024-1164.
- ^ “The size of the BYU 9024 animal”. svpow.com (June 16, 2019). 16 April 2022時点のオリジナルよりアーカイブ。June 16, 2019閲覧。
- ^ Molina-Perez & Larramendi (2020). Dinosaur Facts and Figures: The Sauropods and Other Sauropodomorphs. New Jersey: Princeton University Press. p. 36
- ^ “2億年前の大型恐竜、本当は重くなかった 英米研究”. AFPBB News. AFP. (2012年6月7日) 2017年7月2日閲覧。
- ^ コルバート, モラレス & ミンコフ 2004, pp. 191–194
- ^ コルバート, モラレス & ミンコフ 2004, p. 199
- ^ Theoretical aspects of cursorial adaptations in dinosaurs (Walter P Coombs Jr:1978)
- ^ Dinosauromorph research sheds light on dinosaur evolution(CBC NEWS:2015)
- ^ What, if anything, is a cursor? Categories versus continua for determining locomotor habit in mammals and dinosaurs (MT Carrano:1999)
- ^ ヴォード 2008, p. 249
- ^ 平山廉 2007, pp. 66–67
- ^ 疋田努 2002, p. 72
- ^ 平山廉 2007, p. 70
- ^ , 松岡 & 安藤 2009, p. 11
- ^ , 松岡 & 安藤 2009, p. 21
- ^ フジテレビトリビア普及委員会『トリビアの泉〜へぇの本〜 3』講談社、2003年。
- ^ Vinther, J., Briggs, D. E. G., Prum, R. O. and Saranathan, V. 2008 The colour of fossil feathers.
- ^ “恐竜の色はどのようにして復元するの?|恐竜・古生物 Q&A|FPDM: 福井県立恐竜博物館”. www.dinosaur.pref.fukui.jp. 2022年1月22日閲覧。
- ^ “Yale Scientists First to Reveal Flamboyant Colors of a Dinosaur’s Feathers” (英語). YaleNews (2010年2月4日). 2022年1月22日閲覧。
- ^ a b 鳥類の起源参照
- ^ a b 長谷川政美『系統樹をさかのぼって見えてくる進化の歴史 僕たちの祖先を探す15億年の旅』ベレ出版〈BERET SCIENCE〉、2014年、102ほか頁。ISBN 978-4-86064-410-9。
- ^ 骨「年輪」、草食哺乳類にも=恐竜冷血説を否定-欧州チーム[リンク切れ]
- ^ Meike Köhler, Nekane Marín-Moratalla, Jordana Xavier, Ronny Aanes (2012年7月19日). “Seasonal bone growth and physiology in endotherms shed light on dinosaur physiology”. Nature. Nature Publishing Group. 2017年7月2日閲覧。
- ^ スウィーテク 2011, pp. 170–171
- ^ a b c “恐竜はどうやって巣を作るの?卵はどんなふうに並べるの?”. 福井県立恐竜博物館. 2023年11月16日閲覧。
- ^ “恐竜の繁殖戦略と孵化方法の多様性”. Dino World. 2023年11月16日閲覧。
- ^ Plot R. (1677) The natural history of Oxford-shire : Being an essay toward the natural history of England. Chap. V. Of formed stones.
- ^ 真鍋真 (2005)「人間と恐竜との出会い」. 恐竜博2005-恐竜から鳥への進化. 図録
- ^ Buckland W. (1824) Notice on the Megalosaurus or great fossil lizard. Stonesfield. Transactions of the Geological Society of London 1(2):390-396.
- ^ Mantell, Gideon A. (1825). “Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex”. Philosophical Transactions of the Royal Society 115: 179–186. doi:10.1098/rstl.1825.0010. ISSN 0261-0523. JSTOR 107739.
- ^ von Meyer, H. (1861). "Archaeopteryx litographica (Vogel-Feder) und Pterodactylus von Solenhofen." Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde: 678-679.
- ^ Huxley, Thomas H. (1868). "On the animals which are most nearly intermediate between birds and reptiles". Annals of the Magazine of Natural History 4 (2): 66–75.
- ^ Heilmann, Gerhard (1926). The Origin of Birds. London: Witherby. pp. 208pp
- ^ Paul, G.S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. JHU Press. ISBN 978-0-8018-6763-7
- ^ Padian, Kevin. (2004). “Basal Avialae”. In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. ISBN 978-0-520-24209-8
- ^ Camp, Charles L. (1936). “A new type of small theropod dinosaur from the Navajo Sandstone of Arizona”. Bulletin of the University of California Department of Geological Sciences 24: 39–65.
- ^ Ostrom, John H. (1973). "The ancestry of birds". Nature 242 (5393): 136. Bibcode:1973Natur.242..136O. doi:10.1038/242136a0.
- ^ Gauthier, Jacques. (1986). "Saurischian monophyly and the origin of birds". In Padian, Kevin. (ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8. pp. 1–55.
- ^ Holtz, Jr., T.R. (2000). "Classification and evolution of the dinosaur groups". In Paul, G.S. The Scientific American Book of Dinosaurs. St. Martin's Press. pp. 140–168.
- ^ Burke, A. C. and A. Feduccia (1997). "Developmental patterns and the identification of homologies in the avian hand." Science 278(5338): 666-668.
- ^ Wagner, G. P., and J. Gauthier 1999. 1,2,3 = 2,3,4: a solution to the problem of the homology of the digits in the avian hand. PNAS, 96, 5111-5116.
- ^ Vargas, A. O., T. Kohlsdorf, et al. (2008). "The evolution of HoxD-11 expression in the bird wing: insights from Alligator mississippiensis." PLoS One 3(10): e3325.
- ^ Tamura, K., N. Nomura, et al. (2011). "Embryological evidence identifies wing digits in birds as digits 1, 2, and 3." Science 331(6018): 753-757.
- ^ Xu, X., Clark, J.M., Mo, J., Choiniere, J., Forster, C.A., Erickson, G.M., Hone, D.W.E., Sullivan, C., Eberth, D.A., Nesbitt, S., Zhao, Q., Hernandez, R., Jia, C.-K., Han, F.-L., and Guo, Y. (2009). "A Jurassic ceratosaur from China helps clarify avian digital homologies." Nature, 459(18): 940–944. doi:10.1038/nature08124
- ^ Horner, J. R. and M. B. Goodwin (2008). "Ontogeny of cranial epi-ossifications in Triceratops." Journal of Vertebrate Paleontology 28(1): 134-144.
- ^ Longrich, N. R. and D. J. Field (2012). "Torosaurus is not Triceratops: ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy." PLoS One 7(2): e32623.
- ^ コルバート, モラレス & ミンコフ 2004, p. 190
- ^ MIT’s New Evidence on Dinosaur Evolution(Jennifer Chu:2020)
- ^ 疋田努 2002, p. 43
- ^ 金子隆一『哺乳類型爬虫類 ヒトの知られざる祖先』朝日新聞社〈朝日選書〉、1998年、36頁。ISBN 4022597097。
- ^ Nagao, Takumi (1936-07). “Nipponosaurus Sachalinensis : A New Genus and Species of Trachodont Dinosaur from Japanese Saghalien”. Journal of the Faculty of Science, Hokkaido Imperial University. Ser. 4, Geology and mineralogy (The Hokkaido Imperial University) 3 (2): 185-220. hdl:2115/35768.
- ^ 『北海道中川町の恐竜化石を新属新種「パラリテリジノサウルス・ジャポニクス」と命名~恐竜類テリジノサウルス科の爪の進化~』(プレスリリース)北海道大学、岡山理科大学、中川町エコミュージアムセンター、2022年5月9日。2022年5月9日閲覧。
- ^ 久保田克博「日本産の中生代恐竜化石目録 2022年版」(PDF)『群馬県立自然史博物館研究報告』第27号、群馬県立自然史博物館、2023年3月、157-170頁、CRID 1520859470312165120、ISSN 13424092。
- ^ 宮原れい (2019年9月6日). “通称・むかわ竜、学名が「カムイサウルス・ジャポニクス」に決定 新属新種の恐竜だと正式に認定される”. ねとらぼ. 2021年1月7日閲覧。
- ^ Mortimer, Michael (2008年). “Neotheropoda”. The Theropod Database. 2009年11月17日閲覧。
- ^ “新種化石06「桑島化石壁産出の恐竜化石に学名がつきました」|白山市公式ホームページ”. 白山市公式ホームページ. 2023年3月22日閲覧。
- ^ 「福井の恐竜化石発掘調査30年 きっかけの発見者、松田亜規さん」『毎日新聞』2019年7月3日。2021年2月14日閲覧。
- ^ Kobayashi, Y. and Azuma, Y. (2003). "A new iguanodontian (Dinosauria; Ornithopoda), form the lower Cretaceous Kitadani Formation of Fukui Prefecture, Japan". Journal of Vertebrate Paleontology 23(1): 166-175
- ^ “フクイラプトルという学名になりました”. 福井県立恐竜博物館. 2020年6月27日閲覧。
- ^ Azuma, Y.; Shibata, M. (2010). “Fukuititan nipponensis, a new titanosauriform sauropod from the Early Cretaceous Tetori Group of Fukui Prefecture, Japan”. Acta Geologica Sinica - English Edition 84 (3): 454–462. doi:10.1111/j.1755-6724.2010.00268.x.
- ^ “勝山市の発掘現場から発見された恐竜に新しい学名が付けられました”. 福井県立恐竜博物館 (2015年3月10日). 2017年7月2日閲覧。
- ^ “新恐竜はティラノサウルスもどき 98年から発掘の化石、福井 | 全国のニュース”. 福井新聞ONLINE. 2023年9月8日閲覧。
- ^ “恐竜博物館の調査研究情報、長崎初の大型獣脚類(肉食恐竜)の化石について”. 福井県立恐竜博物館 (2013年7月8日). 2017年7月2日閲覧。
- ^ 共同通信 (2019年1月28日). “国内最古級の恐竜化石発見、熊本 獣脚類の肋骨か - 共同通信 | This Kiji”. 共同通信. 共同通信. 2019年1月29日閲覧。
参考文献
[編集]- David E. Fastovsky、デヴィッド・ワイシャンペル『恐竜学 進化と絶滅の謎』真鍋真(訳)、丸善、2006年。ISBN 978-4-621-07734-4。
- David B. Weishampel, Peter Dodson, Halszka Osmólska, ed (2004). The Dinosauria (second edition ed.). University of California Press. ISBN 978-0520242098
- ピーター・D・ヴォード『恐竜はなぜ鳥に進化したのか』垂水雄二(訳)、文藝春秋、2008年。ISBN 978-4-16-369960-8。
- エドウィン・ハリス・コルバート、マイケル・モラレス、イーライ・C・ミンコフ『脊椎動物の進化』田隅本生(訳)、築地書館、2004年。ISBN 4-8067-1295-7。
- ブライアン・スウィーテク『移行化石の発見』野中香方子(訳)、文藝春秋、2011年。ISBN 978-4-16-373970-0。
- 疋田努『爬虫類の進化』東京大学出版会、2002年。ISBN 4-13-060179-2。
- 平山廉『カメのきた道 甲羅に秘められた2億年の生命進化』日本放送出版協会〈NHKブックス〉、2007年。ISBN 978-4140910955。
- 松岡廣繁、安藤みき子『鳥の骨探』エヌ・ティー・エス、2009年。ISBN 978-4-86043-276-8。
- 伊地知英信『日本にも恐竜がいた』長谷川善和監修、どうぶつ社、1990年。ISBN 4886224156。
関連項目
[編集]「恐竜」に関する情報が検索できます。
- 絶滅した動物一覧 - 恐竜を含む絶滅した全ての動物の一覧。
- 化石爬虫類の一覧 - 恐竜を含む絶滅した爬虫類の一覧。
- 恐竜の一覧 - 主な恐竜の属の一覧。
- 恐竜ルネッサンス
- バーナム・ブラウン - 恐竜に深く関わった人物。
- ギデオン・マンテル - 恐竜に深く関わった人物。
- 恐竜エキスポふくい2000
- ディノサウロイド - 仮説に基づいて想像された「恐竜人間」
- パレオアート - 古代生物の調査で判明した生態などをもとに作られた絵画。
- 恐竜の食餌 - 内容物化石や糞の化石から分析されている食餌の内容について
- 恐竜が登場する映画の一覧 - 恐竜が登場した作品。
外部リンク
[編集]- 恐竜センター (群馬県神流町 恐竜センター)
- 恐竜図鑑 (FPDM 福井県立恐竜博物館)
- 白山恐竜パーク白峰(石川県白山市)
- いわき市石炭化石館(福島県いわき市)
- 人と自然の博物館(兵庫県)
- 恐竜発掘project(日本テレビ系テレビ番組『ザ!鉄腕!DASH!!』)
- 恐竜渓谷ふくい勝山ジオパーク(福井県勝山市)
- ディノ・ネット デジタル恐竜展示室(国立科学博物館、凸版印刷) - 博物館や骨格等をオンラインで確認できるサイト
- 『恐竜』 - コトバンク
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||



| 国立図書館 | |
|---|---|
| その他 | |