
「真核生物」の版間の差分
出典の著者名らしき文字が誤っているように思えたので出典に合わせて修正。(誰も気にしていないのかもしれない) |
英語版 Eukaryote の 2024-03-14T17:51:32(UTC)版と、英語版 Template:Stem group kingdoms の 2024-03-14T17:58:24(UTC)版 を翻訳して記事を強化しました。詳細は ノート:真核生物#報告: 英語版の翻訳による記事の強化 をご覧ください。 タグ: サイズの大幅な増減 ビジュアルエディター |
||
| 1行目: | 1行目: | ||
{{生物分類表 |
{{生物分類表 |
||
| 色 = 原生生物界 |
| 色 = 原生生物界 |
||
| 名称 = 真核生物 |
| 名称<!--name--> = 真核生物<br><span style="font-size: 95%;line-height: 95%;">Eukaryota</span> |
||
| fossil_range = [[スタテリアン]] – [[完新世]] {{long fossil range| |
| fossil_range = [[スタテリアン]] – [[完新世|現在]] {{long fossil range|1650|0 |earliest=2300}} |
||
| 画像 = |
| 画像 = {{Multiple image |
||
| perrow = 2/2/2 |
|||
| 画像キャプション ぬやま |
|||
| total_width = 270 |
|||
| ドメイン = '''真核生物''' {{sname||Eukaryota}} |
|||
| image1 = Rhodomonas salina CCMP 322.jpg |
|||
| caption1 = [[クリプチスタ]]<!-- {{sname||Cryptista}} --> |
|||
| image2 = Ranunculus asiaticus4LEST.jpg |
|||
| caption2 = [[緑色植物亜界]]<!-- {{sname||Viridiplantae}} --> |
|||
| image3 = Trypanosoma sp. PHIL 613 lores.jpg |
|||
| caption3 = [[エクスカバータ#分類|ディスコバ]]<!-- {{sname||Discoba}} --> |
|||
| image4 = Chaos carolinensis Wilson 1900.jpg |
|||
| caption4 = [[アメーボゾア]]<!-- {{sname||Amoebozoa}} --> |
|||
| image5 = Ammonia tepida.jpg |
|||
| caption5 = [[リザリア]]<!-- {{sname||Rhizaria}} --> |
|||
| image6 = Isotricha intestinalis.jpg |
|||
| caption6 = [[アルベオラータ]]<!-- {{sname||Alveolata}} --> |
|||
| image7 = Osmia rufa couple (aka).jpg |
|||
| caption7 = [[動物界]]<!-- {{sname||Animalia}} --> |
|||
| image8 = Boletus edulis (Tillegem).jpg |
|||
| caption8 = [[菌界]]<!-- {{sname||Fungi}} --> |
|||
| border = infobox |
|||
}} |
|||
| ドメイン<!--taxon--> = '''真核生物''' {{sname||Eukaryota}} |
|||
| 学名 = Eukaryota |
| 学名 = Eukaryota |
||
| 学名命名者<!--authority--> = ([[エドゥアール・シャットン]], 1925) [[ロバート・ホイッタカー]] & [[リン・マーギュリス]], 1978 |
|||
| シノニム = |
|||
| 下位分類名<!--subdivision_ranks--> = [[スーパーグループ (分類学)|スーパーグループ]]と[[界 (分類学)|界]]<ref name="AdlBass2019"/> |
|||
* Eukarya |
|||
| 下位分類<!--subdivision--> = |
|||
| 和名 = 真核生物 (しんかくせいぶつ) |
|||
<div style="font-size: 95%;line-height: 95%;"> |
|||
| 英名 = Eukaryote |
|||
| 下位分類名 = 下位分類群 |
|||
| 下位分類 = |
|||
* (和名なし) {{sname||Discoba}} |
|||
* [[ヘミマスティゴフォラ]] {{sname||Hemimastigophora}} |
|||
* [[ディアフォレティケス]] {{sname||Diaphoretickes}} |
* [[ディアフォレティケス]] {{sname||Diaphoretickes}} |
||
** [[アーケプラスチダ|古色素体類]] {{sname||Archaeplastida}} |
|||
** [[ハクロビア]] {{sname||Hacrobia}} |
|||
** [[SARスーパーグループ]] {{sname||SAR supergroup}} |
** [[SARスーパーグループ]] {{sname||SAR supergroup}} |
||
* [[ |
** [[ハプティスタ]] {{sname||Haptista}} |
||
* |
** [[クリプチスタ]] {{sname||Cryptista}} |
||
** [[アーケプラスチダ]] {{sname||Archaeplastida}} ([[植物]]を含む) |
|||
** {{ill2|プロヴォラ|en|Provora}} {{sname||Provora}}<ref name="Provora"/> |
|||
* [[ヘミマスティゴフォラ]] {{sname||Hemimastigophora}} |
|||
* [[メタモナダ]] {{sname||Metamonada}} |
|||
* {{ill2|マラウィモナス|en|Malawimonadida}} {{sname||Malawimonadida}} |
|||
* {{ill2|アンキロモナス|en|Ancyromonadida}} {{sname||Ancyromonadida}} |
|||
* {{ill2|CRuMs|en|CRuMs}} {{sname||CRuMs}} |
|||
* [[アモルフェア]] {{sname||Amorphea}} |
* [[アモルフェア]] {{sname||Amorphea}} |
||
** [[アメーボゾア]] {{sname||Amoebozoa}} |
|||
** [[ブレビアテア]] {{sname||Breviatea}} |
|||
** {{ill2|アプソモナディダ|en|Apusomonadida}} {{sname||Apusomonadida}} |
|||
** [[オピストコンタ]] {{sname||Opisthokonta}} |
|||
*** [[ホロマイコータ]] {{sname||Holomycota}} ([[菌界]]を含む) |
|||
*** [[ホロゾア]] {{sname||Holozoa}} ([[動物]]を含む) |
|||
| シノニム<!--synonyms--> = * Eucarya {{au2|[[カール・ウーズ|ウーズ]]<!-- en:Carl Woese|Woese --> et al. 1990}}<ref name="w1990"/> |
|||
* Eukarya {{au2|[[リン・マーギュリス|マーギュリス]]<!-- en:Lynn Margulis|Margulis--> 1996}}<ref name="Margulis 1996"/> |
|||
| 和名 = 真核生物 (しんかくせいぶつ) |
|||
| 英名 = Eukaryote |
|||
</div> |
|||
{{!}}- style="background:#e0d0b0;" {{!}} |
|||
}} |
}} |
||
'''真核生物'''(しんかくせいぶつ、[[ラテン語|羅]]: {{Sname|Eukaryota}}、[[ |
'''真核生物'''(しんかくせいぶつ、[[ラテン語|羅]]: {{Sname|Eukaryota}}、{{Lang-en-short|eukaryotes}})は、生物学の[[ドメイン (分類学)|ドメイン]] {{Snamei|Eukaryota}} あるいは {{Snamei|Eukarya}} を構成し、[[細胞]]の中に[[細胞核|核]]と呼ばれる[[細胞小器官]]を持つ[[生物]]である。すべての[[動物]]、[[植物]]、[[菌類]]、そして多くの[[単細胞生物]]は真核生物である。真核生物は、[[原核生物]]の2つのグループすなわち[[細菌]]と[[古細菌]]と並び、[[生命|生命体]]の主要なグループを構成している。真核生物は生物の個体数としては少数派であるが、一般的にはるかに大きいので、その集団的な地球規模での[[バイオマス]]は原核生物よりもはるかに大きい。 |
||
真核生物は表面上、[[アスガルド古細菌]]に属する[[古細菌]]が進化して出現したように見える。このことは、生命のドメインは細菌と古細菌の{{Ill2|2ドメイン説|en|Two-domain system|label=2つ}}だけで、真核生物は古細菌の中に組み込まれていることを意味する。真核生物が最初に出現したのは[[古原生代]]のことで、おそらくは[[鞭毛虫|鞭毛のある細胞]]と考えられる。有力な[[進化]]理論によれば、真核生物は、嫌気性のアスガルド古細菌と好気性の[[シュードモナス門]](旧: プロテオバクテリア)の[[細胞内共生説|共生]]によって誕生し、後者から[[ミトコンドリア]]が形成されたという。[[シアノバクテリア]]との共生による2回目のエピソードで、[[葉緑体]]を持つ植物の祖先が誕生した。 |
|||
==概要== |
|||
生物を基本的な[[遺伝]]の仕組みや[[生物化学|生化学]]的性質を元に分類する3[[ドメイン (分類学)|ドメイン]]説では、[[古細菌]]、[[細菌]]と共に生物界を3分する。古細菌および細菌の2ドメインは合わせて[[原核生物]](Prokaryote)とも呼ばれる。真核生物は、原核生物に比べて大型で形態的に多様性に富むという特徴を持つ。かつての[[五界説|5界説]]における[[動物界]]、[[植物界]]、[[菌界]]、[[原生生物界]]の4界はすべて今日では真核生物に含まれる。 |
|||
真核細胞(真核生物の細胞)は、[[細胞核|核]]、[[小胞体]]、[[ゴルジ装置]]などの[[細胞小器官|膜結合細胞小器官]]を持つ。真核生物には[[単細胞]]と[[多細胞]]とがある。それに比べ、原核生物は一般的に単細胞である。単細胞の真核生物は[[原生生物]]と呼ばれることもある。真核生物は[[有糸分裂]]による[[無性生殖]]と、[[減数分裂]]と[[配偶子]]融合([[受精]])による[[有性生殖]]の両方を行うことができる。 |
|||
== 真核細胞の構造 == |
|||
[[ファイル:Biological cell.svg|thumb|350px|left|真核細胞の模式図。1.[[核小体]] 2.[[細胞核]] 3.[[リボソーム]] 4.[[小嚢]] 5.粗面[[小胞体]] 6.[[ゴルジ体]] 7.[[細胞骨格]] 8.滑面[[小胞体]] 9.[[ミトコンドリア]] 10.[[液胞]] 11.[[細胞質基質]] 12.[[リソソーム]] 13.[[中心体]]]] |
|||
真核生物の細胞は一般的に原核生物の細胞よりも大きく、場合によっては1000倍以上の[[体積]]を持つこともある。細胞内にはさまざまな[[細胞小器官]]がある。細胞核は必要な物質のみ透過する穴の開いた二重の膜で覆われており、核液と[[遺伝]]情報を保持する [[デオキシリボ核酸|DNA]] を含んでいる。細胞のその他の部分は[[細胞質]]と呼ばれ、[[細胞骨格]]によって支えられている。 |
|||
__TOC__ |
|||
核の周囲を板状とチューブ状の[[小胞体]]が取り巻いている。チューブ状の物は滑面小胞体、板状の物は粗面小胞体と呼ばれており、粗面小胞体にはいくつもの[[リボソーム]]が張り付き、細胞内での物質の生成、伝達が原核生物と同程度に潤滑させる器官である。リボソーム内で合成された[[蛋白質|タンパク質]]が小胞体に渡され、[[小胞]]に入れられて細胞全体に分配される。ほとんどの真核生物では小胞は[[ゴルジ体]]に蓄積される。小胞には様々な種類があり、これらの動きを合わせて細胞内組織が構成されている。 |
|||
== 多様性 == |
|||
その他にも多くの器官が存在している。原核細胞と異なり、真核細胞の中には異化作用と[[酸素]]の消費に関係する[[ミトコンドリア]]がある。[[植物]]や藻の系列では細胞内に[[光合成]]を行う[[葉緑体]]も含まれている。葉緑体を内部に持つ原核生物も存在しているが、それぞれ別々に細胞内に取り込まれたと考えられている([[細胞内共生説]])。真核生物の多くは細胞表面に[[鞭毛]]や[[繊毛]]があり、移動に使用したり、あるいは受容器官の働きをしている物もある。 |
|||
{{further|生物<!-- Organism-->}} |
|||
== 繁殖 == |
|||
[[細胞分裂]]の際には、まず[[細胞分裂|核分裂]]が行われる。一般に、核内のDNAは細胞分裂に先立って、より集められて[[染色体]]になり、DNAはこの染色体ごとに新しい2つの細胞のために糸状の構造(紡錘糸)によって分かれて運ばれる。分かれた染色体のコピーが渡される。これを有糸分裂と呼ぶ。真核生物の染色体は直鎖状であり、末端には[[テロメア]]と呼ばれる構造がある。 |
|||
真核[[生物]]は、直径3マイクロメートルに満たない[[ピコゾア]]門のような微小な単一[[細胞]]から<ref name="Seenivasan Sausen Medlin Melkonian 2013 p=e59565">{{cite journal |last1=Seenivasan |first1=Ramkumar |last2=Sausen |first2=Nicole |last3=Medlin |first3=Linda K. |last4=Melkonian |first4=Michael |name-list-style=vanc |title=Picomonas judraskeda Gen. Et Sp. Nov.: The First Identified Member of the Picozoa Phylum Nov., a Widespread Group of Picoeukaryotes, Formerly Known as 'Picobiliphytes' |journal=PLOS ONE |volume=8 |issue=3 |date=26 March 2013 |doi=10.1371/journal.pone.0059565 |page=e59565|pmid=23555709 |pmc=3608682 |bibcode=2013PLoSO...859565S |doi-access=free }}</ref>、体重190[[トン]]、体長33.6メートル(110 ft)の[[シロナガスクジラ]]のような[[動物]]<ref name="Wood">{{cite book |last=Wood |first=Gerald |name-list-style=vanc |title=The Guinness Book of Animal Facts and Feats |year=1983 |isbn=978-0-85112-235-9 |url=https://archive.org/details/guinnessbookofan00wood |publisher=Enfield, Middlesex : Guinness Superlatives }}</ref>、あるいは高さ120メートルにもなる(390 ft)の[[セコイア]]のような[[植物]]まで、形態的に多様性に富むさまざまな生物を指す<ref>{{cite web |url=http://www.conifers.org/cu/Sequoia.php |title=Sequoia sempervirens |work=The Gymnosperm Database |editor=Earle CJ |date=2017 |access-date=2017-09-15 |archive-date=2016-04-01 |archive-url=https://web.archive.org/web/20160401041103/http://www.conifers.org/cu/Sequoia.php |url-status=live }}</ref>。多くの真核生物は単細胞生物で、[[原生生物]]と呼ばれる非公式なグループにはこれらの多くが含まれるが、[[ジャイアントケルプ]]({{Snamei|[[:en:Macrocystis|Macrocystis]]}})のような長さ61メートル(200 ft)にもなる多細胞生物もいる<ref name="Hoek 95">{{cite book |last1=van den Hoek |first1=C. |last2=Mann |first2=D.G. |last3=Jahns |first3=H.M. |name-list-style=vanc |year=1995 |url=https://books.google.com/books?id=xuUoiFesSHMC |title=Algae An Introduction to Phycology |publisher=Cambridge University Press |location=Cambridge |isbn=0-521-30419-9 |access-date=7 April 2023 |archive-date=10 February 2023 |archive-url=https://web.archive.org/web/20230210172546/https://books.google.com/books?id=xuUoiFesSHMC |url-status=live }}</ref>。多細胞の真核生物には、動物、植物、[[真菌類]]が含まれるが、やはりこれらのグループにも多くの単細胞[[種 (分類学)|種]]が含まれる<ref name="burki2014" />。真核生物の細胞は通常、[[原核生物]]([[細菌]]や[[古細菌]])よりもはるかに大きく、その体積は約10,000倍である<ref name="DeRennaux 2001 pp. 329–332">{{cite book |last=DeRennaux |first=B. |name-list-style=vanc |title=Encyclopedia of Biodiversity |chapter=Eukaryotes, Origin of |publisher=Elsevier |year=2001 |volume=2 |doi=10.1016/b978-0-12-384719-5.00174-x |pages=329–332|isbn=9780123847201 }}</ref><ref>{{cite journal |title=Deep-sea microorganisms and the origin of the eukaryotic cell |url=http://protistology.jp/journal/jjp47/JJP47YAMAGUCHI.pdf |vauthors=Yamaguchi M, Worman CO |journal=Japanese Journal of Protozoology |volume=47 |number=1,2| date=2014 |pages=29–48 |archive-url=https://web.archive.org/web/20170809103456/http://protistology.jp/journal/jjp47/JJP47YAMAGUCHI.pdf|archive-date=9 August 2017 |url-status=dead}}</ref>。真核生物は[[生物]]の数の中では少数派にすぎないが、その多くがはるかに大きいため、それらの世界全体の[[バイオマス]](468ギガトン)は、原核生物(77ギガトン)よりもはるかに大きく、植物だけで[[地球]]の総バイオマスの81%以上を占めている<ref name="The biomass of Earth">{{Cite journal |last1=Bar-On |first1=Yinon M. |last2=Phillips |first2=Rob |last3=Milo |first3=Ron |date=2018-05-17 |df=dmy-all |title=The biomass distribution on Earth |journal=Proceedings of the National Academy of Sciences |volume=115 |issue=25 |pages=6506–6511 |doi=10.1073/pnas.1711842115 |issn=0027-8424 |pmid=29784790 |pmc=6016768|bibcode=2018PNAS..115.6506B |doi-access=free }}</ref>。 |
|||
ほとんどの真核生物では[[有性生殖]]が行われる。減数分裂後、染色体の半数を2つ合わせて核の合成を行う。これには様々なパターンが存在する。 |
|||
<gallery class="center" mode="nolines" widths="225" heights="225" caption="真核生物の大きさは、単細胞から何トンもある生物までさまざまである"> |
|||
File:Gram-negative Bacteria and Paramecium forming cyst.jpg|(左) [[原核生物]]の[[細菌]](小さな円筒形の細胞)と、(右) 単細胞の真核生物[[ゾウリムシ]]の大きさを比較した顕微鏡像 |
|||
File:California Redwood National Park (216450575).jpeg|[[セコイア]] |
|||
File:Anim1754 - Flickr - NOAA Photo Library (rotated).jpg|[[シロナガスクジラ]] |
|||
</gallery> |
|||
真核生物は多彩な系統であり、主に[[微生物|微細な生物]]から構成されている<ref name="Burki Roger Brown Simpson 2020 pp. 43–55" />。多細胞性は何らかの形で、真核生物の中で少なくとも25回は[[収斂進化|独立して進化]]してきた<ref name="Grosberg2007">{{cite journal |last1=Grosberg |first1=RK |last2=Strathmann |first2=RR |name-list-style=vanc |year=2007 |title=The evolution of multicellularity: A minor major transition? |journal=[[:en:Annu Rev Ecol Evol Syst|Annu Rev Ecol Evol Syst]] |volume=38 |pages=621–654 |doi=10.1146/annurev.ecolsys.36.102403.114735 |url=https://grosberglab.faculty.ucdavis.edu/wp-content/uploads/sites/453/2017/05/2007-Grosberg-R.-K.-and-R.-R.-Strathmann.pdf |access-date=8 April 2023 |archive-date=14 March 2023 |archive-url=https://web.archive.org/web/20230314222721/https://grosberglab.faculty.ucdavis.edu/wp-content/uploads/sites/453/2017/05/2007-Grosberg-R.-K.-and-R.-R.-Strathmann.pdf |url-status=live }}</ref><ref>{{cite journal |last1=Parfrey |first1=L.W. |last2=Lahr |first2=D.J.G. |name-list-style=vanc |year=2013 |title=Multicellularity arose several times in the evolution of eukaryotes |journal=BioEssays |volume=35 |issue=4 |pages=339–347 |doi=10.1002/bies.201200143 |pmid=23315654 |s2cid=13872783 |url=http://www.producao.usp.br/bitstream/handle/BDPI/45022/339_ftp.pdf?sequence=1&isAllowed=y |access-date=8 April 2023 |archive-date=25 July 2014 |archive-url=https://web.archive.org/web/20140725235332/http://www.producao.usp.br/bitstream/handle/BDPI/45022/339_ftp.pdf?sequence=1&isAllowed=y |url-status=live }}</ref>。複雑な多細胞生物は、[[アメーバ属]]の集合体である[[粘菌]]類を除けば、[[動物]]、[[真菌類|真菌]]類、[[褐藻]]類、[[紅藻]]類、[[緑藻]]類、[[陸上植物]]の6つの真核生物の系統の中で進化してきたにすぎない<ref>{{cite journal |last1=Popper |first1=Zoë A. |last2=Michel |first2=Gurvan |last3=Hervé |first3=Cécile |last4=Domozych |first4=David S. |last5=Willats |first5=William G.T. |last6=Tuohy |first6=Maria G. |last7=Kloareg |first7=Bernard |last8=Stengel |first8=Dagmar B. |name-list-style=vanc |year=2011 |title=Evolution and diversity of plant cell walls: From algae to flowering plants |journal=Annual Review of Plant Biology |volume=62 |pages=567–590 |pmid=21351878 |hdl=10379/6762 |hdl-access=free |s2cid=11961888 |doi=10.1146/annurev-arplant-042110-103809}}</ref>。真核生物はゲノムの類似性に基づいてグループ分けされているため、グループには目に見える共通の特徴がないことが多い<ref name="Burki Roger Brown Simpson 2020 pp. 43–55">{{cite journal |last1=Burki |first1=Fabien |last2=Roger |first2=Andrew J. |last3=Brown |first3=Matthew W. |last4=Simpson |first4=Alastair G.B. |name-list-style=vanc |title=The New Tree of Eukaryotes |journal=Trends in Ecology & Evolution |publisher=Elsevier BV |volume=35 |issue=1 |year=2020 |issn=0169-5347 |doi=10.1016/j.tree.2019.08.008 |pages=43–55|pmid=31606140 |s2cid=204545629 |doi-access=free |url=https://uu.diva-portal.org/smash/get/diva2:1387649/FULLTEXT01 }}</ref>。 |
|||
== 起源 == |
|||
かつては[[細菌]]、[[アーキア]] (古細菌)、真核生物の3ドメインは共通の祖先から別個に誕生したとも考えられていたが<ref>{{Cite journal|last=Woese|first=C. R.|date=2002-06-25|title=On the evolution of cells|url=http://www.pnas.org/cgi/doi/10.1073/pnas.132266999|journal=Proceedings of the National Academy of Sciences|volume=99|issue=13|pages=8742–8747|language=en|doi=10.1073/pnas.132266999|issn=0027-8424|pmid=12077305|pmc=124369}}</ref>、現在では古細菌の中から真核生物が進化したとする説が有力になりつつある(かつての'''[[エオサイト説]]'''に近い)<ref name=":1">[[#矢﨑(2020)]] p.72.</ref><ref name=":10">{{Cite journal|last=Zaremba-Niedzwiedzka|first=Katarzyna|last2=Caceres|first2=Eva F.|last3=Saw|first3=Jimmy H.|last4=Bäckström|first4=Disa|last5=Juzokaite|first5=Lina|last6=Vancaester|first6=Emmelien|last7=Seitz|first7=Kiley W.|last8=Anantharaman|first8=Karthik|last9=Starnawski|first9=Piotr|date=2017-01|title=Asgard archaea illuminate the origin of eukaryotic cellular complexity|url=http://www.nature.com/articles/nature21031|journal=Nature|volume=541|issue=7637|pages=353–358|language=en|doi=10.1038/nature21031|issn=0028-0836}}</ref><ref>{{Cite journal|last=Spang|first=Anja|last2=Saw|first2=Jimmy H.|last3=Jørgensen|first3=Steffen L.|last4=Zaremba-Niedzwiedzka|first4=Katarzyna|last5=Martijn|first5=Joran|last6=Lind|first6=Anders E.|last7=van Eijk|first7=Roel|last8=Schleper|first8=Christa|last9=Guy|first9=Lionel|date=2015-05|title=Complex archaea that bridge the gap between prokaryotes and eukaryotes|url=http://www.nature.com/articles/nature14447|journal=Nature|volume=521|issue=7551|pages=173–179|language=en|doi=10.1038/nature14447|issn=0028-0836|pmid=25945739|pmc=4444528}}</ref><ref>{{Cite journal|last=Liu|first=Yang|last2=Makarova|first2=Kira S.|last3=Huang|first3=Wen-Cong|last4=Wolf|first4=Yuri I.|last5=Nikolskaya|first5=Anastasia N.|last6=Zhang|first6=Xinxu|last7=Cai|first7=Mingwei|last8=Zhang|first8=Cui-Jing|last9=Xu|first9=Wei|date=2021-05-27|title=Expanded diversity of Asgard archaea and their relationships with eukaryotes|url=http://www.nature.com/articles/s41586-021-03494-3|journal=Nature|volume=593|issue=7860|pages=553–557|language=en|doi=10.1038/s41586-021-03494-3|issn=0028-0836}}</ref>。特に、[[アスガルド古細菌]]から進化した事が[[分子系統解析]]から示唆されている。アスガルド古細菌は2010年代にその存在が確認され、真核生物と多くの類似性を有することが知られている<ref name=":1" />。 |
|||
== 特徴 == |
|||
{{clade|{{clade |
|||
|label1= |
|||
{{further|細胞#真核細胞<!-- Cell (biology)#Eukaryotic cells-->}} |
|||
|1= [[細菌]] |
|||
|label2= [[古細菌]] |
|||
=== 核 === |
|||
|2={{clade |
|||
|label1=[[アスガルド古細菌]] |
|||
真核生物の決定的な特徴は、その[[細胞]]が[[細胞核|核]]を持っていることである。真核({{Linktext|eukaryote|pref=wikt:en}})という用語は、[[ギリシア語|ギリシャ語]]の{{Linktext|εὖ|pref=wikt:en}}(''eu''、よく、うまく)と{{Linktext|κάρυον|pref=wikt:en}}(''karyon''、仁、核)からその名前がつけられた<ref name="OnlineEtDict">{{OEtymD|eukaryotic}}</ref>。真核細胞には、[[細胞小器官]]と呼ばれるさまざまな内膜結合構造と、細胞の組織と形状を規定する[[細胞骨格]]がある。核は細胞の[[デオキシリボ核酸|DNA]]を保持しており、[[染色体]]と呼ばれる線状の束に分かれている<ref>{{Cite journal |last1=Bonev |first1=B |last2=Cavalli |first2=G |name-list-style=vanc |date=14 October 2016 |title=Organization and function of the 3D genome |journal=Nature Reviews Genetics |volume=17 |issue=11 |pages=661–678 |doi=10.1038/nrg.2016.112 |pmid=27739532 |hdl=2027.42/151884 |s2cid=31259189 |hdl-access=free}}</ref>。これらの染色体は、真核生物に特有の[[有糸分裂]]の過程で核分裂が起こる際、微小管[[紡錘体]]によって2つの同じ集まりに分離される<ref>{{Cite web |title=Chromosome Segregation: The Role of Centromeres |last=O'Connor |first=Clare |work=Nature Education |date= 2008|access-date=18 February 2024 |url= https://www.nature.com/scitable/topicpage/chromosome-segregation-in-mitosis-the-role-of-242/ |quote=eukar }}</ref>。 |
|||
=== 生化学 === |
|||
真核生物は多くの点で原核生物とは異なっており、たとえば、[[ステラン]]合成のような独特な生化学的経路を持っている<ref name="sterane" />。真核生物のそのシグネチャー[[タンパク質]]<!-- signature proteins -->は、他の生命ドメインのタンパク質とは[[相同|相同性]]をもたないが、真核生物の間では普遍的なもののようである。これらのタンパク質には、細胞骨格、複雑な[[転写 (生物学)|転写]]機構、膜選別システム、[[核膜孔]]、および生化学的経路におけるいくつかの[[酵素]]などである<ref name="pmid11805300">{{cite journal |vauthors=Hartman H, Fedorov A |title=The origin of the eukaryotic cell: a genomic investigation |journal=Proceedings of the National Academy of Sciences of the United States of America |volume=99 |issue=3 |pages=1420–5 |date=February 2002 |pmid=11805300 |pmc=122206 |doi=10.1073/pnas.032658599 |bibcode=2002PNAS...99.1420H |doi-access=free }}</ref>。 |
|||
=== 内部膜 === |
|||
{{further|細胞内膜系<!-- Endomembrane system -->}} |
|||
{{multiple image |
|||
| image1 = Prokaryote cell.svg |
|||
| width1 = 5<!--to scale! the point is this is the relative size, thanks-->0 |
|||
| caption1 = <small>[[原核生物]], 同じスケール</small> |
|||
| image2 = Endomembrane system diagram en (edit).svg |
|||
| width2 = 300 |
|||
| caption2 = [[細胞内膜系|内膜系]]を持つ真核細胞 |
|||
| footer = 真核細胞は体積で原核細胞の約10,000倍大きく、[[細胞小器官#種類|膜結合細胞小器官]]を含んでいる |
|||
}} |
|||
真核生物の細胞にはさまざまな膜結合構造があり、それらの集まりが[[細胞内膜系|内膜系]]を形成している<ref>{{cite book |vauthors=Linka M, Weber AP |chapter=Evolutionary Integration of Chloroplast Metabolism with the Metabolic Networks of the Cells |veditors=Burnap RL, Vermaas WF |title=Functional Genomics and Evolution of Photosynthetic Systems |publisher=Springer |year=2011 |isbn=978-94-007-1533-2 |page=215 |chapter-url=https://books.google.com/books?id=WfzEgaLibuwC&pg=PA215 |access-date=27 October 2015 |archive-date=29 May 2016 |archive-url=https://web.archive.org/web/20160529130011/https://books.google.com/books?id=WfzEgaLibuwC&pg=PA215 |url-status=live }}</ref>。[[小胞]]や[[液胞]]と呼ばれる単純な区画は、他の膜からの出芽によって形成される。多くの細胞は、[[エンドサイトーシス]]という過程(外膜が{{Ill2|陥入|en|Invagination}}してからつまみ取るように小胞を形成する)を通じて食物やその他の物質を摂取する<ref>{{cite book| vauthors=Marsh M | title=Endocytosis| publisher=Oxford University Press| year=2001| page=vii| isbn=978-0-19-963851-2}}</ref>。それに対して、[[エキソサイトーシス]]によって小胞から放出される細胞産物もある<ref>{{Cite journal |last1=Stalder |first1=Danièle |last2=Gershlick |first2=David C. |name-list-style=vanc |date=November 2020|title=Direct trafficking pathways from the Golgi apparatus to the plasma membrane |journal=Seminars in Cell & Developmental Biology |volume=107 |pages=112–125 |doi=10.1016/j.semcdb.2020.04.001 |pmc=7152905 |pmid=32317144}}</ref>。 |
|||
核は[[核膜]]と呼ばれる二重膜に囲まれており、[[核膜孔]]が物質の出入りを可能にしている<ref name="TheNuclearEnvelope-NCBI">{{cite journal | vauthors=Hetzer MW | title=The nuclear envelope | journal=Cold Spring Harbor Perspectives in Biology | volume=2 | issue=3 | pages=a000539 | date=March 2010 | pmid=20300205 | pmc=2829960 | doi=10.1101/cshperspect.a000539 }}</ref>。核膜のさまざまな管状や板状の延長部分が[[小胞体]]を形成し、{{Ill2|タンパク質標的化|en|Protein targeting|label=タンパク質の輸送}}と成熟に関与している。[[粗面小胞体]]は、タンパク質を合成する[[リボソーム]]で覆われた小胞体である。生成したタンパク質は内部空間あるいは内腔に入り、その後一般に、[[滑面小胞体]]から出芽した小胞に取り込まれる<ref>{{cite web |title=Endoplasmic Reticulum (Rough and Smooth) |url=http://bscb.org/learning-resources/softcell-e-learning/endoplasmic-reticulum-rough-and-smooth/ |publisher=British Society for Cell Biology |access-date=12 November 2017 |archive-date=24 March 2019 |archive-url=https://web.archive.org/web/20190324055727/https://bscb.org/learning-resources/softcell-e-learning/endoplasmic-reticulum-rough-and-smooth/ |url-status=live }}</ref>。ほとんどの真核生物では、これらのタンパク質を輸送する小胞は放出され、[[ゴルジ装置]]と呼ばれる扁平槽({{Ill2|シスターネ|en|Cisterna}}が積み重なった小器官でさらに修飾される<ref>{{cite web |title=Golgi Apparatus |url=http://bscb.org/learning-resources/softcell-e-learning/golgi-apparatus/ |publisher=British Society for Cell Biology |access-date=12 November 2017 |archive-url=https://web.archive.org/web/20171113060218/http://bscb.org/learning-resources/softcell-e-learning/golgi-apparatus/ |archive-date=13 November 2017 |url-status=dead }}</ref>。 |
|||
小胞は特殊化することもあり、たとえば[[リソソーム]]は、細胞質内の[[生体分子]]を分解する[[消化酵素]]を含んでいる<ref>{{cite web |title=Lysosome |url=http://bscb.org/learning-resources/softcell-e-learning/lysosome/ |publisher=British Society for Cell Biology |access-date=12 November 2017 |archive-url=https://web.archive.org/web/20171113060214/http://bscb.org/learning-resources/softcell-e-learning/lysosome/ |archive-date=13 November 2017 |url-status=dead }}</ref>。 |
|||
=== ミトコンドリア === |
|||
{{main |ミトコンドリア<!-- Mitochondrion -->}} |
|||
[[File:Mitochondrion structure.svg |thumb |upright=1.4 |基本的に真核生物には[[ミトコンドリア]]が存在し、独自に[[デオキシリボ核酸|DNA]]を持つことから原核細胞にも似ている ]] |
|||
[[ミトコンドリア]]は真核細胞に存在する細胞小器官である。ミトコンドリアは、通称「細胞の発電所<ref>{{cite journal |vauthors=Saygin D, Tabib T, Bittar HE, Valenzi E, Sembrat J, Chan SY, Rojas M, Lafyatis R |display-authors=3 |title=Transcriptional profiling of lung cell populations in idiopathic pulmonary arterial hypertension |journal=Pulmonary Circulation |volume=10 |issue=1 |pages=131–144 |date=July 1957 |pmid=32166015 |doi=10.1038/scientificamerican0757-131 |pmc=7052475 |bibcode=1957SciAm.197a.131S }}</ref>」と呼ばれ、糖や脂肪を酸化してエネルギーを貯蔵する[[アデノシン三リン酸]](ATP)[[分子]]を生成し、エネルギーを供給する機能を持つ<ref name="Voet">{{cite book | vauthors=Voet D, Voet JC, Pratt CW |name-list-style=vanc |title=Fundamentals of Biochemistry |edition=2nd |publisher=John Wiley and Sons |year=2006 |pages=[https://archive.org/details/fundamentalsofbi00voet_0/page/547 547, 556] |isbn=978-0471214953 |url=https://archive.org/details/fundamentalsofbi00voet_0/page/547 }}</ref><ref>{{cite web |url=http://www.madsci.org/posts/archives/2006-05/1146679455.Ev.r.html |title=Re: Are there eukaryotic cells without mitochondria? |date=1 May 2006 |work=madsci.org |vauthors=Mack S |access-date=24 April 2014 |archive-date=24 April 2014 |archive-url=https://web.archive.org/web/20140424224700/http://www.madsci.org/posts/archives/2006-05/1146679455.Ev.r.html |url-status=live }}</ref>。ミトコンドリアは、[[リン脂質二重膜]]でできた[[ミトコンドリア#構造|2つの膜]]で覆われ、[[ミトコンドリア内膜|内側の膜]]は[[クリステ]]という折り畳まれた構造になっていて、そこで[[好気呼吸]]が行われる<ref>{{cite journal |last1=Zick |first1=M |last2=Rabl |first2=R |last3=Reichert |first3=AS |name-list-style=vanc |title=Cristae formation-linking ultrastructure and function of mitochondria. |journal=Biochimica et Biophysica Acta (BBA) - Molecular Cell Research |date=January 2009 |volume=1793 |issue=1 |pages=5–19 |pmid=18620004 |doi=10.1016/j.bbamcr.2008.06.013 |doi-access=}}</ref>。 |
|||
ミトコンドリアは[[ミトコンドリアDNA|独自のDNA]]を持ち、そのDNAは起源とする{{Ill2|環状染色体|en|Circular chromosome|label=細菌DNA}}と構造的に類似しており、真核生物のRNAよりも細菌のRNAに近い構造的のRNAを生成する[[rRNA]]と[[tRNA]]の遺伝子をコードしている<ref name="MolecGene">{{cite book |vauthors=Watson J, Hopkins N, Roberts J, Steitz JA, Weiner A |title=Molecular Biology of the Gene |date=1988 |publisher=The Benjamin/Cummings Publishing Company, Inc. |location=Menlo Park, California |isbn=978-0-8053-9614-0 |page=[https://archive.org/details/molecularbiology0004unse/page/1154 1154] |edition=Fourth |chapter=28: The Origins of Life |chapter-url=https://archive.org/details/molecularbiology0004unse/page/1154 }}</ref>。 |
|||
一部の真核生物、たとえば[[メタモナス類]]の[[ジアルジア属]]({{Snamei|Giardia}})や[[トリコモナス]]({{Snamei|Trichomonas}})、[[アメーバ動物門]]の[[ペロミクサ]]({{Snamei|Pelomyxa}})はミトコンドリアを欠いているように見えるが、いずれも[[ハイドロジェノソーム]]や[[マイトソーム]]のようなミトコンドリア由来の細胞小器官を持っており、ミトコンドリアは二次的に失われたものである<ref name="Karn" />。これらは細胞質内の酵素作用によってエネルギーを得ている<ref>{{cite web |url=http://www.iflscience.com/plants-and-animals/first-eukaryote-found-lack-mitochondria |title=Scientists Shocked To Discover Eukaryote With NO Mitochondria |date=13 May 2016 |vauthors=Davis JL |website=IFL Science |access-date=2016-05-13 |archive-url=https://web.archive.org/web/20190217214255/https://www.iflscience.com/plants-and-animals/first-eukaryote-found-lack-mitochondria/ |archive-date=17 February 2019 |url-status=dead }}</ref><ref name="Karn">{{cite journal |vauthors=Karnkowska A, Vacek V, Zubáčová Z, Treitli SC, Petrželková R, Eme L, Novák L, Žárský V, Barlow LD, Herman EK, Soukal P, Hroudová M, Doležal P, Stairs CW, Roger AJ, Eliáš M, Dacks JB, Vlček Č, Hampl V |display-authors=3 |title=A Eukaryote without a Mitochondrial Organelle |journal=Current Biology |volume=26 |issue=10 |pages=1274–1284 |date=May 2016 |pmid=27185558 |doi=10.1016/j.cub.2016.03.053 |doi-access=free }}</ref>。 |
|||
=== プラスチド === |
|||
{{main|プラスチド<!-- Plastid -->}} |
|||
[[File:Chloroplast II.svg|thumb|upright=1.2|[[プラスチド]]の最も一般的な種類は[[葉緑体]]で、葉緑体は[[クロロフィル]]を含み、[[光合成]]によって有機化合物を生成する。 ]] |
|||
植物やさまざまな[[藻類]]は、ミトコンドリアだけでなく[[プラスチド]]も持っている。プラスチドは、ミトコンドリアと同様に{{Ill2|葉緑体DNA|en|Chloroplast DNA|label=独自のDNA}}を持ち、[[内生生物|内部共生]](この場合は[[シアノバクテリア]])から進化した。それらは通常、[[葉緑体]]の形を取り、シアノバクテリアのように[[クロロフィル]]を含み、[[光合成]]によって[[グルコース]]などの有機化合物を生成する。また、栄養素の貯蔵<!-- food storage -->を担うものもある。プラスチドはおそらく単一の起源を持つが、すべてのプラスチドを持つグループが密接に関連しているわけではない。それどころか、真核生物の中には、{{Ill2|二次内部共生|en|Secondary endosymbiosis|label=二次的な内部共生}}あるいは摂取によって、他の生物からそれらを獲得したものもある<ref>{{cite book | vauthors=Sato N |year=2006 |pages= 75–102 |title=The Structure and Function of Plastids |volume=23 |veditors=Wise RR, Hoober JK |publisher= Springer Netherlands|chapter=Origin and Evolution of Plastids: Genomic View on the Unification and Diversity of Plastids |isbn=978-1-4020-4060-3 |doi=10.1007/978-1-4020-4061-0_4 |series=Advances in Photosynthesis and Respiration}}</ref>。光合成細胞や葉緑体の捕獲と隔離、すなわち[[盗葉緑体現象|盗葉緑体化]]は、現代の多くの種類の真核生物で見られる<ref name="pmid18518896">{{cite journal |vauthors=Minnhagen S, Carvalho WF, Salomon PS, Janson S |title=Chloroplast DNA content in Dinophysis (Dinophyceae) from different cell cycle stages is consistent with kleptoplasty |journal=Environ. Microbiol. |volume=10 |issue=9 |pages=2411–7 |date=September 2008 |pmid=18518896 |doi=10.1111/j.1462-2920.2008.01666.x |bibcode=2008EnvMi..10.2411M }}</ref><ref name="Kleptoplastidy Hypothesis">{{cite journal | vauthors=Bodył A | s2cid=24613863 | title=Did some red alga-derived plastids evolve via kleptoplastidy? A hypothesis | journal=Biological Reviews of the Cambridge Philosophical Society | volume=93 | issue=1 | pages=201–222 | date=February 2018 | pmid=28544184 | doi=10.1111/brv.12340 }}</ref>。 |
|||
=== 細胞骨格 === |
|||
{{main|細胞骨格<!-- Cytoskeleton -->}} |
|||
[[File:FluorescentCells.jpg|thumb|顕微鏡下で観察したウシ肺動脈の内皮細胞の細胞骨格。細胞核は青、微小管は緑、フィラメント状アクチンは赤で標識されている。 ]] |
|||
[[細胞骨格]]は、細胞が動いたり、形を変化させたり、物質の輸送を可能にするモーター構造のために、剛構造と結合点を提供する。モーター構造は[[アクチン]]の[[マイクロフィラメント]](微小線維)であり、α-{{Ill2|アクチニン|en|Actinin}}、[[フィンブリン]]、[[フィラミン]]などの[[アクチン結合タンパク質]]が膜下の[[細胞皮質|皮質]]や繊維束<!-- submembranous cortical layers and bundles -->に存在する。微小管の[[モータータンパク質]]、[[ダイニン]]と[[キネシン]]、そしてアクチンフィラメントの[[ミオシン]]が、ネットワークに動的な特性を与える<ref>{{Cite book |chapter=Molecular Motors |title=Molecular Biology of the Cell |edition=4th |url=https://www.ncbi.nlm.nih.gov/books/NBK26888/ |date=2002-01-01 |first1=Bruce |last1=Alberts |first2=Alexander |last2=Johnson |first3=Julian |last3=Lewis |first4=Martin |last4=Raff |first5=Keith |last5=Roberts |first6=Peter |last6=Walter |name-list-style=vanc |publisher=Garland Science |location=New York |isbn=978-0-8153-3218-3 |access-date=6 April 2023 |archive-date=8 March 2019 |archive-url=https://web.archive.org/web/20190308094109/https://www.ncbi.nlm.nih.gov/books/NBK26888/ |url-status=live }}</ref><ref name="Sweeney">{{cite journal |vauthors=Sweeney HL, Holzbaur EL |title=Motor Proteins |journal=Cold Spring Harbor Perspectives in Biology |volume=10 |issue=5 |pages= a021931|date=May 2018 |pmid=29716949 |pmc=5932582 |doi=10.1101/cshperspect.a021931 |url=}}</ref>。 |
|||
多くの真核生物は、[[鞭毛]]と呼ばれる細長い運動性の細胞質突起、あるいは[[繊毛]]と呼ばれる多数の短い構造を持っている。これらの{{Ill2|波動毛|en|Undulipodium|label=細胞小器官}}は、運動、摂食、感覚などさまざまに関与している。それらは主に[[チューブリン]]から構成され、原核生物の鞭毛とはまったく異なる。これらは[[中心小体]]から生成する[[微小管]]の束によって支えられており、2本の1本鎖を9本の2本鎖が取り囲むように配列しているのが特徴である。鞭毛は、[[ストラメノパイル]]({{Snamei|Stramenopiles}})の多くに見られるように、管状小毛({{Ill2|マスチゴネマ|en|Mastigoneme}})を持つこともある。それらの内部は[[細胞質]]と連続している<ref name="Bardy Ng Jarrell 2003">{{cite journal | vauthors=Bardy SL, Ng SY, Jarrell KF | title=Prokaryotic motility structures | journal=Microbiology | volume=149 | issue=Pt 2 | pages=295–304 | date=February 2003 | pmid=12624192 | doi=10.1099/mic.0.25948-0 | doi-access=free }}</ref><ref name="Silflow Lefebvre 2001">{{cite journal | vauthors=Silflow CD, Lefebvre PA | title=Assembly and motility of eukaryotic cilia and flagella. Lessons from Chlamydomonas reinhardtii | journal=Plant Physiology | volume=127 | issue=4 | pages=1500–7 | date=December 2001 | pmid=11743094 | pmc=1540183 | doi=10.1104/pp.010807 }}</ref>。 |
|||
中心小体は、鞭毛を持たない細胞や細胞群でもよく存在するが、[[針葉樹類]]や[[顕花植物]]にはどちらもない。これらは一般に、さまざまな微小管性鞭毛根<!-- microtubular roots -->を生じさせるグループに存在する。これらは細胞骨格の主要な構成要素を形成し、しばしば数回の細胞分裂の過程で組み立てられ、一方の鞭毛は親から受け継ぎ、もう一方はそこから派生する。中心小体は核分裂の際に紡錘体の形成に関与する<ref>{{cite book |vauthors=Vorobjev IA, Nadezhdina ES |title=The centrosome and its role in the organization of microtubules |volume=106 |pages=227–293 |year=1987 |pmid=3294718 |doi=10.1016/S0074-7696(08)61714-3 |isbn=978-0-12-364506-7 |series=International Review of Cytology }}</ref>。 |
|||
=== 細胞壁 === |
|||
{{main|細胞壁<!-- Cell wall -->}} |
|||
植物、藻類、真菌類、そしてほとんどの[[クロムアルベオラータ]]類の細胞は細胞壁に囲まれているが、動物の細胞は細胞壁に囲まれていない。これは[[細胞膜]]の外側にある層で、細胞を構造的に支え、保護し、濾過機構を提供する。細胞壁はまた、水が細胞内に侵入したときの{{Ill2|細胞溶解|en|Cytolysis|label=過膨張}}を防ぐ役割も果たす<ref name="Howland 2000">{{cite book | vauthors=Howland JL | year=2000 | title=The Surprising Archaea: Discovering Another Domain of Life | pages=69–71 | publisher=Oxford University Press | location=Oxford | isbn=978-0-19-511183-5}}</ref>。 |
|||
[[陸上植物]]の一次細胞壁を構成する主な[[多糖|多糖類]]は、[[セルロース]]、[[ヘミセルロース]]、[[ペクチン]]である。セルロース・{{Ill2|ミクロフィブリル|en|Microfibril}}はヘミセルロースと結合し、ペクチン・マトリックスに埋め込まれている。一次細胞壁で最も一般的なヘミセルロースは[[キシログルカン]]である<ref name="Fry1989">{{cite journal | vauthors=Fry SC |title=The Structure and Functions of Xyloglucan |journal=Journal of Experimental Botany |volume=40 |issue=1 |year=1989 |pages=1–11 |doi=10.1093/jxb/40.1.1}}</ref>。 |
|||
=== 有性生殖 === |
|||
{{further|{{ill2|有性生殖の進化|en|Evolution of sexual reproduction}}}} |
|||
[[File:Sexual cycle N-2N.svg|thumb|[[有性生殖]]には、細胞内に[[染色体]]が一つずつ存在する[[単数体|単相]]と、二本ずつ存在する[[二倍体|複相]]を交互に繰り返す[[生活環]]がなくてはならない。真核生物では、[[減数分裂]]によって単数体の[[配偶子]]が作られ、2つの配偶子が融合して二倍体の[[受精卵|接合子]]が形成される。]] |
|||
真核生物は[[有性生殖]]を伴う[[生活環]]を持ち、各細胞に染色体が1つずつしか存在しない[[単数体|単相]]と、各細胞に染色体が2つずつ存在する[[二倍体|複相]]とを交互に繰り返す。複相は、[[卵細胞|卵子]]と[[精子]]などの2つの配偶子が融合して、[[受精卵|接合子]]を形成することで成立する。この接合子は、[[有糸分裂]]によって細胞分裂を繰り返しながら成体に成長し、ある段階で染色体数を減らして{{Ill2|遺伝的変動|en|Genetic variability}}を生み出す[[減数分裂]]によって単数体配偶子を形成する<ref>{{cite book |last=Hamilton |first=Matthew B. |name-list-style=vanc |title=Population genetics |url=https://archive.org/details/populationgeneti00hami |url-access=limited |year=2009 |publisher=[[:en:Wiley-Blackwell|Wiley-Blackwell]] |isbn=978-1-4051-3277-0 |page=[https://archive.org/details/populationgeneti00hami/page/n69 55]}}</ref>。この様式にはかなりの多様性がある。植物には{{Ill2|世代交代|en|Alternation of generations|label=単数体と二倍体の両方の多細胞期|preserve=1}}がある<ref>{{cite journal |last1=Taylor |first1=TN |last2=Kerp |first2=H |last3=Hass |first3=H |name-list-style=vanc |year=2005 |title=Life history biology of early land plants: Deciphering the gametophyte phase |journal=Proceedings of the National Academy of Sciences of the United States of America |volume=102 |issue=16 |pages=5892–5897 |doi=10.1073/pnas.0501985102 |pmid=15809414 |pmc=556298 |doi-access=free }}</ref>。真核生物は原核生物よりも代謝率が低く、世代時間が長くなるが、これは真核生物が原核生物よりもはるかに大きく、体積に対する表面積の比が小さいからである<ref name="Lane2011">{{cite journal |vauthors=Lane N |authorlink=Nick Lane |title=Energetics and genetics across the prokaryote-eukaryote divide |journal=Biology Direct |volume=6 |issue=1 |page=35 |date=June 2011 |pmid=21714941 |pmc=3152533 |doi=10.1186/1745-6150-6-35 |doi-access=free }}</ref>。 |
|||
{{ill2|有性生殖の進化|en|Evolution of sexual reproduction}}は、真核生物の原初的な特徴という可能性がある。[[分子系統解析|系統学的分析]]に基づき、Dacksと[[:en:Andrew J. Roger|Roger]]は通性性(facultative sex)がこのグループの共通祖先に存在したと提唱している<ref>{{cite journal |vauthors=Dacks J, Roger AJ |s2cid=9441768 |title=The first sexual lineage and the relevance of facultative sex |journal=Journal of Molecular Evolution |volume=48 |issue=6 |pages=779–783 |date=June 1999 |pmid=10229582 |doi=10.1007/PL00013156 |bibcode=1999JMolE..48..779D }}</ref>。膣トリコモナス({{Snamei|[[:en:Trichomonas vaginalis|Trichomonas vaginalis]]}})と腸鞭毛虫({{Snamei|[[:en:Giardia intestinalis|Giardia intestinalis]]}}、あるいはランブル鞭毛虫({{Snamei|[[:en:Giardia duodenalis|Giardia duodenalis]]}}))という、以前は無性であると考えられていた2つの生物には、減数分裂で機能するコア遺伝子セットが存在する<ref name="Ramesh">{{cite journal |vauthors=Ramesh MA, Malik SB, Logsdon JM |title=A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis |journal=Current Biology |volume=15 |issue=2 |pages=185–191 |date=January 2005 |pmid=15668177 |doi=10.1016/j.cub.2005.01.003 |s2cid=17013247 |doi-access=free }}</ref><ref name="Malik">{{cite journal |vauthors=Malik SB, Pightling AW, Stefaniak LM, Schurko AM, Logsdon JM |title=An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis |journal=PLOS ONE |volume=3 |issue=8 |pages=e2879 |date=August 2007 |pmid=18663385 |pmc=2488364 |doi=10.1371/journal.pone.0002879 |veditors=Hahn MW |bibcode=2008PLoSO...3.2879M |doi-access=free }}</ref>。この2種は、真核生物の進化系統樹の初期に分岐した系統の子孫であることから、コア減数分裂遺伝子、ひいては性が真核生物の共通祖先に存在した可能性がある<ref name="Ramesh" /><ref name="Malik" />。[[リーシュマニア]]({{Snamei|Leishmania}})寄生虫など、かつては無性であると考えられていた種にも性周期がある<ref>{{cite journal |vauthors=Akopyants NS, Kimblin N, Secundino N, Patrick R, Peters N, Lawyer P, Dobson DE, Beverley SM, Sacks DL |title=Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector |journal=Science |volume=324 |issue=5924 |pages=265–268 |date=April 2009 |pmid=19359589 |pmc=2729066 |doi=10.1126/science.1169464 |bibcode=2009Sci...324..265A }}</ref>。以前は無性生物と考えられていたアメーバは、古代は有性生物であり、現在の無性群体<!--asexual groups-->は最近進化した可能性が高い<ref>{{cite journal |vauthors=Lahr DJ, Parfrey LW, Mitchell EA, Katz LA, Lara E |title=The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms |journal=Proceedings: Biological Sciences |volume=278 |issue=1715 |pages=2081–2090 |date=July 2011 |pmid=21429931 |pmc=3107637 |doi=10.1098/rspb.2011.0289 }}</ref>。 |
|||
== 進化 == |
|||
[[File:Openly available illustrations as tools to describe eukaryotic microbial diversity - Journal.pbio.3002395.g001.tif|thumb|主要なサブグループと各グループの代表的なメンバーの簡略図が示された真核生物の系統樹。最近 (2023年現在) の系統発生学的な再構築に基づいて更新された<ref>{{Cite Q|Q123558544|doi-access=free}}</ref>。]] |
|||
=== 分類の歴史 === |
|||
{{further|分類学#分類学の歴史<!-- History of taxonomy-->|リンネ式階層分類体系}}[[古代]]、[[アリストテレス]]や[[テオプラストス]]は、[[動物]]と[[植物]]という2つの生物の系統を識別していた。これらの系統は、18世紀に[[カール・フォン・リンネ|リンネ]]によって[[界 (分類学)|界]](Kingdom)という分類学的な[[階級 (生物学)|階級]]が与えられた。リンネは、[[真菌類]]を植物に含めることに若干の条件をつけたが、後に、真菌類はまったく別個のもので、独立した界を持つに値することがわかった<ref name="Moore1980">{{cite journal |vauthors=Moore RT |year=1980 |title=Taxonomic proposals for the classification of marine yeasts and other yeast-like fungi including the smuts |journal=Botanica Marina |volume=23 |pages=361–373}}</ref>。さまざまな単細胞の真核生物が知られるようになった当初、それらは植物や動物と一緒にされていた。1818年、ドイツの生物学者{{Ill2|ゲオルク・アウグスト・ゴルトフス|en|Georg August Goldfuss|label=ゲオルク・A・ゴルトフス}}は、[[繊毛虫]]のような生物を指すために[[原生動物]]({{Snamei|Protozoa}})という言葉を作り<ref>{{cite journal |author=Goldfuß |title=Ueber die Classification der Zoophyten |journal=Isis, Oder, Encyclopädische Zeitung von Oken |date=1818 |volume=2 |issue=6 |pages=1008–1019 |url=https://www.biodiversitylibrary.org/item/47614#page/530/mode/1up |trans-title=On the classification of zoophytes |language=de |access-date=15 March 2019 |archive-date=24 March 2019 |archive-url=https://web.archive.org/web/20190324105702/https://www.biodiversitylibrary.org/item/47614#page/530/mode/1up |url-status=live }} From p. 1008: ''"Erste Klasse. Urthiere. Protozoa."'' (First class. Primordial animals. Protozoa.) [Note: each column of each page of this journal is numbered; there are two columns per page.]</ref>、このグループは、1866年に[[エルンスト・ヘッケル]]がすべての単細胞真核生物を包括する界、[[原生生物界]]({{Snamei|Protista}})を作るまで拡張された<ref name="Scamardella1999">{{cite journal |vauthors=Scamardella JM |title=Not plants or animals: a brief history of the origin of Kingdoms Protozoa, Protista and Protoctista |year=1999 |journal=[[:en:International Microbiology|International Microbiology]] |volume=2 |issue=4 |pages=207–221 |pmid=10943416 |url=http://www.im.microbios.org/08december99/03%20Scamardella.pdf |url-status=dead |archive-url=https://web.archive.org/web/20110614000656/http://www.im.microbios.org/08december99/03%20Scamardella.pdf |archive-date=14 June 2011 }}</ref><ref name="Rothschild1989">{{cite journal |vauthors=Rothschild LJ |title=Protozoa, Protista, Protoctista: what's in a name? |journal=Journal of the History of Biology |volume=22 |issue=2 |pages=277–305 |year=1989 |pmid=11542176 |doi=10.1007/BF00139515 |s2cid=32462158 |authorlink=Lynn J. Rothschild |url=https://zenodo.org/record/1232387 |access-date=4 February 2020 |archive-date=4 February 2020 |archive-url=https://web.archive.org/web/20200204233203/https://zenodo.org/record/1232387 |url-status=live }}</ref><ref name="Whittaker">{{cite journal |vauthors=Whittaker RH |title=New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms |journal=Science |volume=163 |issue=3863 |pages=150–60 |date=January 1969 |pmid=5762760 |doi=10.1126/science.163.3863.150 |citeseerx=10.1.1.403.5430 |bibcode=1969Sci...163..150W }}</ref>。こうして真核生物は4つの界に分類された。 |
|||
* [[原生生物]]界(Kingdom {{Snamei|Protista}}) |
|||
* [[植物]]界(Kingdom {{Snamei|Plantae}}) |
|||
* [[菌類|菌]]界(Kingdom {{Snamei|Fungi}}) |
|||
* [[動物]]界(Kingdom {{Snamei|Animalia}}) |
|||
当時、原生生物は「原始的な形態」であり、原始的な単細胞の性質が合併した{{Ill2|進化グレード|en|Evolutionary grade|label=進化の一段階}}であると考えられていた<ref name="Rothschild1989" />。 |
|||
19世紀にはすでに、[[細胞核|核]]という構造の有無が生物の分類にとって重要な差異であることは認識されていた。エルンスト・ヘッケルは、細菌などのなんの構造も持たない生物を原生生物の中の[[モネラ]]として区別し、後にシアノバクテリアをここに含めている<ref name="Sapp2005">{{cite journal|author=Sapp, J.|year=2005|title=The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology|journal=Microbiol. Mol. Biol. Rev.|volume=69|issue=2|pages=292-305|doi=10.1128/mmbr.69.2.292-305.2005}}</ref>。しかし当時は[[動物]]と[[植物]]という差異がまず先に立っており、モネラとそれ以外という差異が注目されることはなかった。 |
|||
真核生物という言葉は、文献上[[エドゥアール・シャットン]]が1925年の論文で初めて用いた<ref name="Katscher2004">{{cite journal|author=Katscher, F.|year=2004|title=The History of the Terms Prokaryotes and Eukaryotes| journal=Protist|volume=155|pages=257-263|doi=10.1078/143446104774199637}}</ref>。この論文は{{snamei|Pansporella perplexa}}の分類学的位置を議論するもので、末尾の原生生物の分類表と樹形図の中で{{lang|fr|Eucaryotes}}と{{lang|fr|Procaryotes}}が示されているものの、他には何の説明もなかった<ref>{{cite journal|author=Chatton, E.|year=1925|title=Pansporella perplexa, amoebien à spores protégées parasite de daphnies|journal=Ann. Sci. Nat. Zool.|volume=8|pages=5–84|url=https://gallica.bnf.fr/ark:/12148/bpt6k55415433/f8.item}}</ref>。シャットンの弟子で後に[[ノーベル生理学・医学賞]]を受賞した[[アンドレ・ルヴォフ]]の1932年のモノグラフの冒頭には、シャットンを引用しながら原生生物を原核生物と真核生物に二分する旨の記述がある。ここでは、原核的原生生物を[[細胞核]]や[[ミトコンドリア]]がないもの、真核的原生生物を両者を持つものとしている<ref>{{cite book|author=Lwoff, A.|year=1932|title=Recherches Biochimiques sur la Nutrition des Protozoaires. Le Pouvoir de Synthèse.|publisher=Masson|locastion=Paris}}</ref>。以後、20世紀前半に[[英語]]、[[ドイツ語]]、[[フランス語]]の文献で何度か言及されてはいるが、生物を真核生物と原核生物に二分する方法は一般的な認識とは程遠かった<ref name="Katscher2004" />。たとえば{{仮リンク|ハーバート・コープランド|en|Herbert Copeland}}は1938年に細胞核がない生物を[[モネラ界]]としたが、細胞核がある生物についてはヘッケルの3界(動物界、植物界、原生生物界)をそのまま採用している<ref>{{cite journal|author=Copeland, H.|year=1938|title=The kingdoms of organisms|journal=Quart. Rev. Biol.|volume=13|issue=4|pages=383-420|doi=10.1086/394568}}</ref>。この二分法を普及させたのは、カナダ人の[[細菌学者]] {{Ill2|ロジャー・スタニエ|en|Roger Stanier}}である。彼は1960年から翌年にかけてサバティカルで[[パスツール研究所]]に滞在し、ルヴォフとの議論の中でシャットンの二分法を知り、1962年の論文<ref>{{cite journal|author=Stanier & van Niel|year=1962|title=The concept of a bacterium|url=https://fire.biol.wwu.edu/cmoyer/zztemp_fire/biol497_F13/papers/Stanier_archmicro62.pdf|journal=Arch. Mikrobiol.|volume=42|pages=17–35|doi=10.1007/BF00425185}}</ref>で広く知らしめたのである<ref name="Sapp2005" /><ref name="Katscher2004" />。[[電子顕微鏡]]による[[微細構造]]観察が当たり前のように行われる時代になって、この二分法は広く受け入れられるようになった。 |
|||
{{Ill2|生命の樹 (生物学)|en|Tree of life (biology)|label=生命の樹}}([[系統樹]]も参照)における最古の分岐の理解は、DNAの[[塩基配列]]の決定によって初めて実質的に進展し、1990年に[[カール・ウーズ]]、{{Ill2|オットー・カントラー|en|Otto Kandler}}、{{Ill2|マーク・ウィーリス|en|Mark Wheelis}}らが提唱した最上位の階級を(界ではなく)[[ドメイン (分類学)|ドメイン]]とする体系({{Ill2|3ドメイン系|en|Three-domain system}})が導かれた。彼らは、すべての真核生物の界を「Eucarya」ドメインに統合したが、「真核生物は今後も受け入れられる一般的な同義語である」と述べている<ref name="w1990">{{cite journal |vauthors=Woese CR, Kandler O, Wheelis ML |author1-link=Carl Woese |author2-link=Otto Kandler |author3-link=Mark Wheelis |title=Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya |journal=Proceedings of the National Academy of Sciences of the United States of America |volume=87 |issue=12 |pages=4576–4579 |date=June 1990 |pmid=2112744 |pmc=54159 |doi=10.1073/pnas.87.12.4576 |bibcode=1990PNAS...87.4576W |doi-access=free }}</ref><ref name="Knoll 1992">{{cite journal |last=Knoll |first=Andrew H. |name-list-style=vanc |authorlink=Andrew H. Knoll |title=The Early Evolution of Eukaryotes: A Geological Perspective |journal=Science |volume=256 |issue=5057 |year=1992 |doi=10.1126/science.1585174 |pages=622–627 |pmid=1585174 |bibcode=1992Sci...256..622K |quote=Eucarya, or eukaryotes}}</ref>。1996年、進化生物学者の[[リン・マーギュリス]]は、界とドメインを「包括的」な名前に置き換えて「共生に基づく系統発生」を作ることを提案し、「真核生物(共生由来の有核生物)」という説明を加えた<ref name="Margulis 1996">{{cite journal |last=Margulis |first=Lynn |authorlink=Lynn Margulis |name-list-style=vanc |title=Archaeal-eubacterial mergers in the origin of Eukarya: phylogenetic classification of life |journal=Proceedings of the National Academy of Sciences |volume=93 |issue=3 |date=6 February 1996 |doi=10.1073/pnas.93.3.1071 |pages=1071–1076 |pmid=8577716 |pmc=40032 |bibcode=1996PNAS...93.1071M |doi-access=free }}</ref>。 |
|||
しかしながら、真核生物以外のすべての生物の総称として、原核生物という言葉は今日でも学術論文で用いられている。一方で21世紀に入ると、真核生物は古細菌から派生して出現した系統であるという理解が普及し、生物界を細菌とそれ以外で分ける、上記とは異なる意味合いでの二分法が出現している<ref name=":10">{{Cite journal|last=Zaremba-Niedzwiedzka|first=Katarzyna|last2=Caceres|first2=Eva F.|last3=Saw|first3=Jimmy H.|last4=Bäckström|first4=Disa|last5=Juzokaite|first5=Lina|last6=Vancaester|first6=Emmelien|last7=Seitz|first7=Kiley W.|last8=Anantharaman|first8=Karthik|last9=Starnawski|first9=Piotr|date=2017-01|title=Asgard archaea illuminate the origin of eukaryotic cellular complexity|url=http://www.nature.com/articles/nature21031|journal=Nature|volume=541|issue=7637|pages=353–358|language=en|doi=10.1038/nature21031|issn=0028-0836}}</ref>。 |
|||
=== 系統発生 === |
|||
2014年までに、過去20年間の系統学的研究から、大まかな合意が生まれはじめた<ref name="burki2014">{{cite journal |vauthors=Burki F |title=The eukaryotic tree of life from a global phylogenomic perspective |journal=Cold Spring Harbor Perspectives in Biology |volume=6 |issue=5 |pages=a016147 |date=May 2014 |pmid=24789819 |doi=10.1101/cshperspect.a016147 |pmc=3996474 }}</ref><ref name="UntanglingEarlyEukaryotes">{{cite journal |vauthors=Burki F, Kaplan M, Tikhonenkov DV, Zlatogursky V, Minh BQ, Radaykina LV, Smirnov A, Mylnikov AP, Keeling PJ |display-authors=3 |title=Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista |journal=Proceedings: Biological Sciences |volume=283 |issue=1823 |pages=20152802 |date=January 2016 |pmid=26817772 |pmc=4795036 |doi=10.1098/rspb.2015.2802 }}</ref>。真核生物の大部分は、アモルフェア({{Snamei|Amorphea}}、[[ユニコント]]仮説に似た構成)と、植物とほとんどの藻類系統が含まれる{{Snamei|[[:en:Diphoda|Diphoda]]}}(旧: [[バイコンタ]])と呼ばれる2つの大きなクレードのいずれかに分類される。第3の主要グループである[[エクスカバータ]]({{Snamei|Excavata}})は、[[側系統群|側系統]]であるため、正式な群としては放棄された<ref name="AdlBass2019">{{cite journal |vauthors=Adl SM, Bass D, Lane CE, Lukeš J, Schoch CL, Smirnov A, Agatha S, Berney C, Brown MW, Burki F, Cárdenas P, Čepička I, Chistyakova L, Del Campo J, Dunthorn M, Edvardsen B, Eglit Y, Guillou L, Hampl V, Heiss AA, Hoppenrath M, James TY, Karnkowska A, Karpov S, Kim E, Kolisko M, Kudryavtsev A, Lahr DJ, Lara E, Le Gall L, Lynn DH, Mann DG, Massana R, Mitchell EA, Morrow C, Park JS, Pawlowski JW, Powell MJ, Richter DJ, Rueckert S, Shadwick L, Shimano S, Spiegel FW, Torruella G, Youssef N, Zlatogursky V, Zhang Q |display-authors=3 |title=Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes |journal=The Journal of Eukaryotic Microbiology |volume=66 |issue=1 |pages=4–119 |date=January 2019 |pmid=30257078 |pmc=6492006 |doi=10.1111/jeu.12691 }}</ref>。以下の提案された系統発生図には、エクスカバータの1つの群(ディスコバ、{{Snamei|Discoba}})のみが含まれ<ref name="Brown 2018">{{Cite journal |last1=Brown |first1=Matthew W. |last2=Heiss |first2=Aaron A. |last3=Kamikawa |first3=Ryoma |last4=Inagaki |first4=Yuji |last5=Yabuki |first5=Akinori |last6=Tice |first6=Alexander K |last7=Shiratori |first7=Takashi |last8=Ishida |first8=Ken-Ichiro |last9=Hashimoto |first9=Tetsuo |last10=Simpson |first10=Alastair |last11=Roger |first11=Andrew |name-list-style=vanc |date=2018-01-19 |title=Phylogenomics Places Orphan Protistan Lineages in a Novel Eukaryotic Super-Group|journal=Genome Biology and Evolution |volume=10 |issue=2 |pages=427–433 |doi=10.1093/gbe/evy014 |pmc=5793813|pmid=29360967}}</ref>、[[ピコゾア門|ピコゾア]]門({{Snamei|Picozoa}})は[[紅藻]]({{Snamei|Rhodophyta}})の近縁種であるという2021年の提案が取り入れられている<ref name="SchönZlatogursky2021">{{cite journal |vauthors=Schön ME, Zlatogursky VV, Singh RP, Poirier C, Wilken S, Mathur V, Strassert JF, Pinhassi J, Worden AZ, Keeling PJ, Ettema TJ |display-authors=3 |title=Picozoa are archaeplastids without plastid |journal=Nature Communications |year=2021 |volume=12 |issue=1 |page=6651 |doi=10.1038/s41467-021-26918-0 |pmid=34789758 |pmc=8599508 |biorxiv=10.1101/2021.04.14.439778 |s2cid=233328713 |url=http://urn.kb.se/resolve?urn=urn:nbn:se:umu:diva-189959 |access-date=20 December 2021 |archive-date=2 February 2024 |archive-url=https://web.archive.org/web/20240202091441/https://umu.diva-portal.org/smash/record.jsf?pid=diva2%3A1614928&dswid=-3028 |url-status=live }}</ref>。プロヴォラ({{Snamei|[[:en:Provora|Provora]]}})は2022年に発見された微生物捕食者の群である<ref name="Provora">{{cite journal |vauthors=Tikhonenkov DV, Mikhailov KV, Gawryluk RM, Belyaev AO, Mathur V, Karpov SA, Zagumyonnyi DG, Borodina AS, Prokina KI, Mylnikov AP, Aleoshin VV, Keeling PJ |display-authors=3 |title=Microbial predators form a new supergroup of eukaryotes |journal=Nature |date=December 2022 |volume=612 |issue=7941 |pages=714–719 |pmid=36477531 |doi=10.1038/s41586-022-05511-5 |bibcode=2022Natur.612..714T |s2cid=254436650 }}</ref>。 |
|||
<!-- 以下の系統発生図は、英語版Wikipediaのテンプレート [[:en:Special:Permalink/1213711338|英語版 Template:Stem group kingdoms の 2024-03-14T17:58:24(UTC)版]] を翻訳して作成した。--> |
|||
{{Clade|label1='''[[真核生物]]/{{small|Eukaryotes}}'''|sublabel1=2200 mya|style1=font-size:90%; line-height:90% |
|||
|{{clade |
|||
|1={{clade |
|||
|1=アンキロモナス [[File:Ancyromonas.png|30px]]<br />{{small|{{sname||Ancyromonadida}}}} |
|||
|2={{clade |
|||
|1={{ill2|マラウィモナス|en|Malawimonad}} [[File:Malawimonas.jpg|30px]]<br />{{small|{{sname||Malawimonad}}a}} |
|||
|2={{clade |
|||
|1=クルムス [[File:Collodictyon pseudopodoa (extracted).jpg|30px]]<br />{{small|{{sname||CRuMs}}}} |
|||
|label2=[[アモルフェア]]<br />{{small|{{sname||Amorphea}}}} |sublabel2=1500 mya |
|||
|2={{clade |
|||
|1=[[アメーボゾア]] [[File:Chaos carolinensis Wilson 1900.jpg|33 px]]<br />{{small|{{sname||Amoebozoa}}}} |
|||
|label2=オバゾア<br >{{small|{{sname||Obazoa}}}} |
|||
|2={{clade |
|||
|1=[[ブレビアテア]] [[File:Mastigamoeba invertens (extracted).jpg|50px]]<br />{{small|{{sname||Breviatea}}}} |
|||
|2={{clade |
|||
|1=アプソモナス [[File:Podomonas kaiyoae C.jpg|30 px]]<br />{{small|{{sname||Apusomonadida}}}} |
|||
|label2=[[オピストコンタ]]<br />{{small|{{sname||Opisthokonta}}}} |
|||
|sublabel2=1300 mya |
|||
|2={{clade |
|||
|1= [[ホロマイコータ]] (真菌を含む) [[File:Asco1013.jpg|30 px]]<br />{{small|{{sname||Holomycota}}}} |
|||
|2= [[ホロゾア]] (動物を含む) [[File:Comb jelly.jpg|33 px]]<br />{{small|{{sname||Holozoa}}}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
|label2={{sname||Diphoda}} |
|||
|sublabel2=Bikonts |
|||
|2={{clade |
|||
|1={{clade |
|||
|1=? [[メタモナダ]] [[File:Giardia lamblia.jpg|30px]]<br />{{small|{{sname||Metamonada}}}} |
|||
|2=ディスコバ [[File:Euglena mutabilis - 400x - 1 (10388739803) (cropped).jpg|28px]]<br />{{small|{{sname||Discoba}}}} |
|||
}} |
|||
|label2=[[ディアフォレティケス]]<br />{{small|{{sname||Diaphoretickes}}}} |
|||
|2={{clade |
|||
|1={{clade |
|||
|1=[[クリプチスタ]] [[File:Rhodomonas salina CCMP 322.jpg|35px]]<br />{{small|{{sname||Cryptista}}}} |
|||
|label2=[[アーケプラスチダ]]<br />{{small|{{sname||Archaeplastida}}}} |
|||
|sublabel2= 1600 mya |
|||
|2={{clade |
|||
|1={{clade |
|1={{clade |
||
| |
|1=[[紅藻]] [[File:Bangia.jpg|20 px]]<br />{{small|{{sname||Rhodophyta}}}} |
||
|2=[[ピコゾア]] [[File:Picomonas judraskeda (SEM).png|23 px]]<br />{{small|{{sname||Picozoa}}}} |
|||
|2=その他のアスガルド古細菌([[側系統]]) |
|||
}} |
|||
| |
|2={{clade |
||
| |
|sublabel1= 1100 mya |
||
|1=[[灰色藻]] [[File:Glaucocystis sp.jpg|30 px]]<br />{{small|{{sname||Glaucophyta}}}} |
|||
|sublabel2= 1000 mya |
|||
|2=[[緑色植物亜界]] (植物を含む) [[File:Pediastrum (cropped).jpg|30 px]]<br />{{small|{{sname||Viridiplantae}}}} |
|||
}} |
|||
}} |
}} |
||
}} |
|||
|2={{clade |
|||
|1=[[ヘミマスティゴフォラ]] [[File:Hemimastix amphikineta.png|20 px]]<br />{{small|{{sname||Hemimastigophora}}}} |
|||
|2={{clade |
|||
|1={{ill2|プロヴォラ|en|Provora}} [[File:Outline drawing of Ubysseya fretuma.svg|25 px]]<br />{{sname||Provora}} |
|||
|2={{clade |
|||
|1=[[ハプチスタ]] [[File:Acanthocystis labeled Picture1.1.png|30 px]]<br />{{small|{{sname||Haptista}}}} |
|||
|label2={{sname||TSAR}} |
|||
|2={{clade |
|||
|1=[[テロネマ門]] [[File:Telonema rivulare (electron micrography).jpg|30 px]]<br />{{small|{{sname||Telonemia}}}} |
|||
|label2=[[SARスーパーグループ|SAR]]<br />{{small|{{sname||SAR supergroup}}}} |
|||
|2={{clade |
|||
|sublabel1= 550 mya |
|||
|1=[[リザリア]] [[File:Ammonia tepida.jpg|30 px]]<br />{{small|{{sname||Rhizaria}}}} |
|||
|label2={{sname||Halvaria}} |
|||
|2={{clade |
|||
|1=[[アルベオラータ]] [[File:Ceratium furca.jpg|40 px]]<br />{{small|{{sname||Alveolata}}}} |
|||
|2=[[ストラメノパイル]] [[File:Ochromonas.png|20 px]] [[File:Gemeiner Blasentang.jpg|30 px]]<br />{{small|{{sname||Stramenopiles}}}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
この系統発生図は上界<!-- great kingdoms -->とその[[ステムグループ]]の一つの見方を示す<ref name="Brown 2018">{{Cite journal |last1=Brown |first1=Matthew W. |last2=Heiss |first2=Aaron A. |last3=Kamikawa |first3=Ryoma |last4=Inagaki |first4=Yuji |last5=Yabuki |first5=Akinori |last6=Tice |first6=Alexander K |last7=Shiratori |first7=Takashi |last8=Ishida |first8=Ken-Ichiro |last9=Hashimoto |first9=Tetsuo |last10=Simpson |first10=Alastair |last11=Roger |first11=Andrew |name-list-style=vanc |date=2018-01-19 |title=Phylogenomics Places Orphan Protistan Lineages in a Novel Eukaryotic Super-Group|journal=Genome Biology and Evolution |volume=10 |issue=2 |pages=427–433 |doi=10.1093/gbe/evy014 |pmc=5793813|pmid=29360967}}</ref><ref name="Picozoa 2021">{{cite journal |vauthors=Schön ME, Zlatogursky VV, Singh RP, Poirier C, Wilken S, Mathur V, Strassert JF, Pinhassi J, Worden AZ, Keeling PJ, Ettema TJ |display-authors=3 |title=Picozoa are archaeplastids without plastid |journal=Nature Communications |year=2021 |volume=12 |issue=1 |page=6651 |doi=10.1038/s41467-021-26918-0 |pmid=34789758 |pmc=8599508 |biorxiv=10.1101/2021.04.14.439778 |s2cid=233328713 |url=http://urn.kb.se/resolve?urn=urn:nbn:se:umu:diva-189959 }}</ref><ref name="Provora 2022">{{cite journal |vauthors=Tikhonenkov DV, Mikhailov KV, Gawryluk RM, Belyaev AO, Mathur V, Karpov SA, Zagumyonnyi DG, Borodina AS, Prokina KI, Mylnikov AP, Aleoshin VV, Keeling PJ |display-authors=3 |title=Microbial predators form a new supergroup of eukaryotes |journal=Nature |date=December 2022 |volume=612 |issue=7941 |pages=714–719 |pmid=36477531 |doi=10.1038/s41586-022-05511-5 |s2cid=254436650 }}</ref><ref name="Burki Roger Brown Simpson 2020 pp. 43–55">{{cite journal |last1=Burki |first1=Fabien |last2=Roger |first2=Andrew J. |last3=Brown |first3=Matthew W. |last4=Simpson |first4=Alastair G.B. |name-list-style=vanc |title=The New Tree of Eukaryotes |journal=Trends in Ecology & Evolution |publisher=Elsevier BV |volume=35 |issue=1 |year=2020 |issn=0169-5347 |doi=10.1016/j.tree.2019.08.008 |pages=43–55|pmid=31606140 |s2cid=204545629 |doi-access=free |url=https://uu.diva-portal.org/smash/get/diva2:1387649/FULLTEXT01 }}</ref>。[[メタモナーダ]]は位置づけが困難で、{{Ill2|ディスコバ|en|Discoba}}あるいは{{Ill2|マラウィモナス|en|Malawimonada}}の姉妹という可能性がある<ref name="Burki Roger Brown Simpson 2020 pp. 43–55"/>。 |
|||
{{clear}} |
|||
=== 真核生物の起源 === |
|||
真核生物には、実際のところ古細菌だけでなく、細菌など他の生物由来の要素も多く含まれている<ref>{{Cite journal|last=Brueckner|first=Julia|last2=Martin|first2=William F|editor-last=Pisani|editor-first=Davide|date=2020-04-01|title=Bacterial Genes Outnumber Archaeal Genes in Eukaryotic Genomes|url=https://academic.oup.com/gbe/article/12/4/282/5788535|journal=Genome Biology and Evolution|volume=12|issue=4|pages=282–292|language=en|doi=10.1093/gbe/evaa047|issn=1759-6653|pmid=32142116|pmc=7151554}}</ref>。真核生物は少なくとも2種以上の生物が合体して誕生したことがほぼ定説となっており、例えばミトコンドリアは、真核生物の母体となった古細菌に[[プロテオバクテリア#アルファプロテオバクテリア綱|αプロテオバクテリア]]が[[細胞内共生説|細胞内共生]]を経て取り込まれて変化したものであるとする説が有力である<ref>{{Cite journal|last=Sagan|first=Lynn|date=1967-03|title=On the origin of mitosing cells|url=https://linkinghub.elsevier.com/retrieve/pii/0022519367900793|journal=Journal of Theoretical Biology|volume=14|issue=3|pages=225–IN6|language=en|doi=10.1016/0022-5193(67)90079-3}}</ref>。ミトコンドリアと同様に、葉緑体はシアノバクテリアが細胞内共生をへて真核生物に取り込みまれたものであると考えられている。ただし葉緑体については、すでに真核生物の誕生後、[[アーケプラスチダ|藻類]]の祖先となる系統でのみ個別に生じた現象であると考えられており、真核生物自体の起源とは関係しない。 |
|||
{{main|{{Ill2|真核生物の最終共通祖先|en|Eukaryogenesis}}<!-- Eukaryogenesis -->}} |
|||
古細菌から真核生物への具体的な道筋はいまだ未解明であり、水素仮説<ref>{{Cite journal|last=Martin|first=William|last2=Müller|first2=Miklós|date=1998-03|title=The hydrogen hypothesis for the first eukaryote|url=http://www.nature.com/articles/32096|journal=Nature|volume=392|issue=6671|pages=37–41|language=en|doi=10.1038/32096|issn=0028-0836}}</ref>、リバース・フローモデル<ref>{{Cite journal|last=Spang|first=Anja|last2=Stairs|first2=Courtney W.|last3=Dombrowski|first3=Nina|last4=Eme|first4=Laura|last5=Lombard|first5=Jonathan|last6=Caceres|first6=Eva F.|last7=Greening|first7=Chris|last8=Baker|first8=Brett J.|last9=Ettema|first9=Thijs J. G.|date=2019-07|title=Proposal of the reverse flow model for the origin of the eukaryotic cell based on comparative analyses of Asgard archaeal metabolism|url=http://www.nature.com/articles/s41564-019-0406-9|journal=Nature Microbiology|volume=4|issue=7|pages=1138–1148|language=en|doi=10.1038/s41564-019-0406-9|issn=2058-5276}}</ref>、E<sup>3</sup>モデル<ref>{{Cite journal|last=Imachi|first=Hiroyuki|last2=Nobu|first2=Masaru K.|last3=Nakahara|first3=Nozomi|last4=Morono|first4=Yuki|last5=Ogawara|first5=Miyuki|last6=Takaki|first6=Yoshihiro|last7=Takano|first7=Yoshinori|last8=Uematsu|first8=Katsuyuki|last9=Ikuta|first9=Tetsuro|date=2020-01-23|title=Isolation of an archaeon at the prokaryote–eukaryote interface|url=http://www.nature.com/articles/s41586-019-1916-6|journal=Nature|volume=577|issue=7791|pages=519–525|language=en|doi=10.1038/s41586-019-1916-6|issn=0028-0836|pmid=31942073|pmc=7015854}}</ref> など多くの仮説が提唱されている<ref>{{Cite journal|last=López-García|first=Purificación|last2=Moreira|first2=David|date=2020-05|title=The Syntrophy hypothesis for the origin of eukaryotes revisited|url=http://www.nature.com/articles/s41564-020-0710-4|journal=Nature Microbiology|volume=5|issue=5|pages=655–667|language=en|doi=10.1038/s41564-020-0710-4|issn=2058-5276}}</ref>。ほとんどの仮説が、古細菌がバクテリアを取り込んだと考えているのに対して、シントロピー・モデル<ref>{{Cite journal|last=Moreira|first=David|last2=López-García|first2=Purificación|date=1998-11|title=Symbiosis Between Methanogenic Archaea and δ-Proteobacteria as the Origin of Eukaryotes: The Syntrophic Hypothesis|url=http://link.springer.com/10.1007/PL00006408|journal=Journal of Molecular Evolution|volume=47|issue=5|pages=517–530|language=en|doi=10.1007/PL00006408|issn=0022-2844}}</ref> と呼ばれる仮説のみ、バクテリア(特に[[プロテオバクテリア|デルタプロテオバクテリア]])が古細菌を取り込んだと推定しており、共生の関係性が他の説とは逆である。この説ではミトコンドリアは古細菌とは別個に取り込まれて成立したとされる。上記の説以外にも、真核生物の細胞核に類似の器官をもつ一部のバクテリア(例えば[[プランクトミケス門|プランクトミケス]])が、真核生物の起源に関与しているとする説も存在する<ref>{{Cite journal|last=Fuerst|first=John A.|last2=Sagulenko|first2=Evgeny|date=2011-06|title=Beyond the bacterium: planctomycetes challenge our concepts of microbial structure and function|url=http://www.nature.com/articles/nrmicro2578|journal=Nature Reviews Microbiology|volume=9|issue=6|pages=403–413|language=en|doi=10.1038/nrmicro2578|issn=1740-1526}}</ref>。 |
|||
[[File:Symbiogenesis 2 mergers.svg|thumb|upright=1.5|[[細胞内共生説]]では、[[古細菌]]と好気性細菌が融合して好気性[[ミトコンドリア]]を持つ真核生物が誕生し、2度目の融合で[[葉緑体]]が加わって[[緑色植物亜界|緑色植物]]が誕生した<ref name=latorre/>]] |
|||
=== 成立年代の推定 === |
|||
真核生物の成立年代は未確定ではあるものの、例えば真核生物に不可欠ないくつかの器官(例えば[[ミトコンドリア]]や、[[ステロール]]を含む[[細胞膜]])<ref>{{Cite journal|last=Roger|first=Andrew J.|last2=Muñoz-Gómez|first2=Sergio A.|last3=Kamikawa|first3=Ryoma|date=2017-11|title=The Origin and Diversification of Mitochondria|url=https://doi.org/10.1016/j.cub.2017.09.015|journal=Current Biology|volume=27|issue=21|pages=R1177–R1192|doi=10.1016/j.cub.2017.09.015|issn=0960-9822}}</ref><ref name=":9">{{Cite journal|last=Hoshino|first=Yosuke|last2=Gaucher|first2=Eric A.|date=2021-06-22|title=Evolution of bacterial steroid biosynthesis and its impact on eukaryogenesis|url=https://www.pnas.org/content/118/25/e2101276118|journal=Proceedings of the National Academy of Sciences|volume=118|issue=25|language=en|doi=10.1073/pnas.2101276118|issn=0027-8424|pmid=34131078|pmc=8237579}}</ref>の成立に酸素が必須なことから、真核生物は24億年前の大酸化イベント以後、[[好気呼吸|好気性]]条件下でおおまかに19億年前頃([[原生代]])には成立したとする説が有力である<ref name=":8"/>。一方で、真核生物は酸素が大気中に含まれていなかった大酸化イベント([[:en:Great_Oxidation_Event|Great Oxidation Event]]; GOE)以前の生活スタイル([[嫌気呼吸]])も保持しており<ref>{{Cite journal|last=Müller|first=Miklós|last2=Mentel|first2=Marek|last3=van Hellemond|first3=Jaap J.|last4=Henze|first4=Katrin|last5=Woehle|first5=Christian|last6=Gould|first6=Sven B.|last7=Yu|first7=Re-Young|last8=van der Giezen|first8=Mark|last9=Tielens|first9=Aloysius G. M.|date=2012-06|title=Biochemistry and Evolution of Anaerobic Energy Metabolism in Eukaryotes|url=https://journals.asm.org/doi/10.1128/MMBR.05024-11|journal=Microbiology and Molecular Biology Reviews|volume=76|issue=2|pages=444–495|language=en|doi=10.1128/MMBR.05024-11|issn=1092-2172|pmid=22688819|pmc=3372258}}</ref><ref>{{Cite book|title=Mitochondria and Anaerobic Energy Metabolism in Eukaryotes: Biochemistry and Evolution|url=https://www.degruyter.com/document/doi/10.1515/9783110612417/html|publisher=De Gruyter|date=2020-12-07|isbn=978-3-11-061241-7|doi=10.1515/9783110612417|first=William F.|last=Martin|first2=Aloysius G. M.|last2=Tielens|first3=Marek|last3=Mentel}}</ref>、最初に誕生した真核生物は[[通性嫌気性生物]]であったと想定される。ちなみに大酸化イベント以前([[太古代]])の地球にもごく少量の酸素は存在していた可能性がある<ref>{{Cite journal|last=Catling|first=David C.|last2=Zahnle|first2=Kevin J.|date=2020-02|title=The Archean atmosphere|url=https://advances.sciencemag.org/lookup/doi/10.1126/sciadv.aax1420|journal=Science Advances|volume=6|issue=9|pages=eaax1420|language=en|doi=10.1126/sciadv.aax1420|issn=2375-2548|pmid=32133393|pmc=7043912}}</ref>。ただ、真核生物を含め好気性生物が太古代にすでに存在していたかについては、それを明確に支持する証拠は現在のところない。最も古い真核生物の痕跡として、27億年前の地層から検出された[[ステラン]]と呼ばれる真核生物由来の有機物質が一時期議論されていたが<ref>{{Cite journal|last=Brocks|first=J. J.|date=1999-08-13|title=Archean Molecular Fossils and the Early Rise of Eukaryotes|url=https://www.sciencemag.org/lookup/doi/10.1126/science.285.5430.1033|journal=Science|volume=285|issue=5430|pages=1033–1036|doi=10.1126/science.285.5430.1033}}</ref><ref>{{Cite journal|last=Waldbauer|first=Jacob R.|last2=Sherman|first2=Laura S.|last3=Sumner|first3=Dawn Y.|last4=Summons|first4=Roger E.|date=2009-03|title=Late Archean molecular fossils from the Transvaal Supergroup record the antiquity of microbial diversity and aerobiosis|url=https://linkinghub.elsevier.com/retrieve/pii/S0301926808002507|journal=Precambrian Research|volume=169|issue=1-4|pages=28–47|language=en|doi=10.1016/j.precamres.2008.10.011}}</ref><ref>{{Cite journal|last=Brocks|first=Jochen J|last2=Buick|first2=Roger|last3=Summons|first3=Roger E|last4=Logan|first4=Graham A|date=2003-11|title=A reconstruction of Archean biological diversity based on molecular fossils from the 2.78 to 2.45 billion-year-old Mount Bruce Supergroup, Hamersley Basin, Western Australia|url=https://linkinghub.elsevier.com/retrieve/pii/S0016703703002096|journal=Geochimica et Cosmochimica Acta|volume=67|issue=22|pages=4321–4335|language=en|doi=10.1016/S0016-7037(03)00209-6}}</ref>、その後これらのステランは当時のものではなく後世の混入であると結論づけられた<ref>{{Cite journal|last=French|first=Katherine L.|last2=Hallmann|first2=Christian|last3=Hope|first3=Janet M.|last4=Schoon|first4=Petra L.|last5=Zumberge|first5=J. Alex|last6=Hoshino|first6=Yosuke|last7=Peters|first7=Carl A.|last8=George|first8=Simon C.|last9=Love|first9=Gordon D.|date=2015-05-12|title=Reappraisal of hydrocarbon biomarkers in Archean rocks|url=https://www.pnas.org/content/112/19/5915|journal=Proceedings of the National Academy of Sciences|volume=112|issue=19|pages=5915–5920|doi=10.1073/pnas.1419563112|pmid=25918387|pmc=4434754}}</ref><ref>{{Cite journal|last=Rasmussen|first=Birger|last2=Fletcher|first2=Ian R.|last3=Brocks|first3=Jochen J.|last4=Kilburn|first4=Matt R.|date=2008-10|title=Reassessing the first appearance of eukaryotes and cyanobacteria|url=http://www.nature.com/articles/nature07381|journal=Nature|volume=455|issue=7216|pages=1101–1104|language=en|doi=10.1038/nature07381|issn=0028-0836}}</ref>。ステランは、真核生物が特徴的に生成する[[ステロール]]が地層中で化石化したものである。現在、真核生物由来のステランとして認められた最も古いものは約8億年前の[[新原生代]]のものにとどまる<ref>{{Cite journal|last=Hoshino|first=Yosuke|last2=Poshibaeva|first2=Aleksandra|last3=Meredith|first3=William|last4=Snape|first4=Colin|last5=Poshibaev|first5=Vladimir|last6=Versteegh|first6=Gerard J. M.|last7=Kuznetsov|first7=Nikolay|last8=Leider|first8=Arne|last9=van Maldegem|first9=Lennart|date=2017-09|title=Cryogenian evolution of stigmasteroid biosynthesis|url=https://advances.sciencemag.org/lookup/doi/10.1126/sciadv.1700887|journal=Science Advances|volume=3|issue=9|pages=e1700887|language=en|doi=10.1126/sciadv.1700887|issn=2375-2548|pmc=5606710|pmid=28948220}}</ref><ref>{{Cite journal|last=Brocks|first=Jochen J.|last2=Jarrett|first2=Amber J. M.|last3=Sirantoine|first3=Eva|last4=Hallmann|first4=Christian|last5=Hoshino|first5=Yosuke|last6=Liyanage|first6=Tharika|date=2017-08|title=The rise of algae in Cryogenian oceans and the emergence of animals|url=https://www.nature.com/articles/nature23457|journal=Nature|volume=548|issue=7669|pages=578–581|language=en|doi=10.1038/nature23457|issn=1476-4687}}</ref>。 |
|||
すべての複雑な細胞とほぼすべての多細胞生物が真核生物に含まれることから、真核生物の起源、すなわち真核発生(''eukaryogenesis'')は、生命の進化における画期的な出来事であった。{{Ill2|真核生物の最終共通祖先|en|Eukaryogenesis}}(LECA)とは、現生するすべての真核生物の起源と仮定されるもので<ref name="Gabaldón">{{cite journal |vauthors=Gabaldón T |title=Origin and Early Evolution of the Eukaryotic Cell |journal=Annual Review of Microbiology |volume=75 |issue=1 |pages=631–647 |date=October 2021 |pmid=34343017 |doi=10.1146/annurev-micro-090817-062213 |s2cid=236916203 }}</ref>、単一の個体ではなく[[個体群|生物学的な集団]]であった可能性が高い<ref name="O'Malley Leger Wideman Ruiz-Trillo pp. 338–344">{{cite journal |vauthors=O'Malley MA, Leger MM, Wideman JG, Ruiz-Trillo I |title=Concepts of the last eukaryotic common ancestor |journal=Nature Ecology & Evolution |volume=3 |issue=3 |pages=338–344 |date=March 2019 |pmid=30778187 |doi=10.1038/s41559-019-0796-3 |bibcode=2019NatEE...3..338O |hdl-access=free |s2cid=67790751 |hdl=10261/201794 }}</ref>。LECAは、核に加え、少なくとも1つの[[中心小体]]と[[鞭毛]]、通性好気性ミトコンドリア、性([[減数分裂]]と[[受精|異型配偶子融合]])、[[キチン]]または[[セルロース]]の細胞壁を持つ休眠[[嚢胞]]<!-- dormant cyst -->、そして[[ペルオキシソーム]]を持つ原生生物であったと考えられている<ref>{{cite journal |vauthors=Leander BS |title=Predatory protists |journal=Current Biology |volume=30 |issue=10 |pages=R510–R516 |date=May 2020 |pmid=32428491 |doi=10.1016/j.cub.2020.03.052 |s2cid=218710816 |doi-access=free }}</ref><ref name="RedAlgalDerivedPlastids">{{cite journal |vauthors=Strassert JF, Irisarri I, Williams TA, Burki F |title=A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids |journal=Nature Communications |volume=12 |issue=1 |pages=1879 |date=March 2021 |pmid=33767194 |pmc=7994803 |doi=10.1038/s41467-021-22044-z |bibcode=2021NatCo..12.1879S |doi-access=free }}</ref><ref name="Koumandou Wickstead Ginger van der Giezen 2013">{{cite journal |last1=Koumandou |first1=V. Lila |last2=Wickstead |first2=Bill |last3=Ginger |first3=Michael L. |last4=van der Giezen |first4=Mark |last5=Dacks |first5=Joel B. |last6=Field |first6=Mark C. |name-list-style=vanc |title=Molecular paleontology and complexity in the last eukaryotic common ancestor |journal=Critical Reviews in Biochemistry and Molecular Biology |volume=48 |issue=4 |year=2013 |doi=10.3109/10409238.2013.821444 |pages=373–396|pmid=23895660 |pmc=3791482 }}</ref>。 |
|||
新原生代以前の真核生物の有無および実態については詳しくわかっていない。2023年、現生の真核生物がもつステロールとは化学構造がやや異なる、”より原始的な”プロトステロールが化石化したものが新原生代以前の地層に広く分布していることが発表され、これらのステロールは現生の真核生物(クラウン・グループ)以前に存在していたステム・グループに属する生物が作り出していた可能性が指摘された<ref>{{Cite journal|last=Brocks|first=Jochen J.|last2=Nettersheim|first2=Benjamin J.|last3=Adam|first3=Pierre|last4=Schaeffer|first4=Philippe|last5=Jarrett|first5=Amber J. M.|last6=Güneli|first6=Nur|last7=Liyanage|first7=Tharika|last8=van Maldegem|first8=Lennart M.|last9=Hallmann|first9=Christian|date=2023-06-22|title=Lost world of complex life and the late rise of the eukaryotic crown|url=https://www.nature.com/articles/s41586-023-06170-w|journal=Nature|volume=618|issue=7966|pages=767–773|language=en|doi=10.1038/s41586-023-06170-w|issn=0028-0836}}</ref>。この説に従えば、現存する{{仮リンク|真核生物発生|label=真核生物の最終共通祖先|en|Eukaryogenesis}}は新原生代まで出現しなかったことになり、それまでは真核生物の前駆段階にあたる何らかの好気性生物が長く繁栄していたことになる。一方で、プロトステロールを含めてステロール自体は細菌が究極的な起源である可能性も指摘されており<ref name=":9" />、新原生代以前のステロール(プロトステロール)を合成していた生物が何者だったのかによって、真核生物の成立過程についての理解は今後大きく変化する可能性がある。 |
|||
運動性の[[嫌気性生物|嫌気性]]古細菌と好気性[[アルファプロテオバクテリア綱|アルファプロテオバクテリア]]綱の[[シンビオジェネシス|内共生併合]]によって、[[ミトコンドリア]]持つLECAそしてすべての真核生物が誕生した。さらにその後、シアノバクテリアとの2回目の内共生により、[[葉緑体]]を持つ植物の祖先が誕生した<ref name="latorre">{{cite book |vauthors=Latorre A, Durban A, Moya A, Pereto J |chapter-url=https://books.google.com/books?id=m3oFebknu1cC&pg=PA326 |chapter=The role of symbiosis in eukaryotic evolution |title=Origins and Evolution of Life: An astrobiological perspective |veditors=Gargaud M, López-Garcìa P, Martin H |year=2011 |location=Cambridge |publisher=Cambridge University Press |pages=326–339 |isbn=978-0-521-76131-4 |access-date=27 August 2017 |archive-date=24 March 2019 |archive-url=https://web.archive.org/web/20190324055723/https://books.google.com/books?id=m3oFebknu1cC&pg=PA326 |url-status=live }}</ref>。 |
|||
ステラン以外の真核生物の痕跡としては、真核生物由来とされる[[微化石]]が21億年前の地層から発見されている<ref>{{Cite journal|last=Albani|first=Abderrazak El|last2=Bengtson|first2=Stefan|last3=Canfield|first3=Donald E.|last4=Bekker|first4=Andrey|last5=Macchiarelli|first5=Roberto|last6=Mazurier|first6=Arnaud|last7=Hammarlund|first7=Emma U.|last8=Boulvais|first8=Philippe|last9=Dupuy|first9=Jean-Jacques|date=2010-07|title=Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago|url=https://www.nature.com/articles/nature09166|journal=Nature|volume=466|issue=7302|pages=100–104|language=en|doi=10.1038/nature09166|issn=1476-4687}}</ref>。ただし、これらの化石が真に真核生物由来かどうかはなお議論の必要がある。19億年前の地層から見つかった、コイル状の多細胞生物と推定されるGrypaniaは真核生物として一定の支持を得ている最古の化石の一つである<ref name=":8">{{Cite journal|last=Knoll|first=A.h|last2=Javaux|first2=E.j|last3=Hewitt|first3=D|last4=Cohen|first4=P|date=2006-06-29|title=Eukaryotic organisms in Proterozoic oceans|url=https://royalsocietypublishing.org/doi/10.1098/rstb.2006.1843|journal=Philosophical Transactions of the Royal Society B: Biological Sciences|volume=361|issue=1470|pages=1023–1038|doi=10.1098/rstb.2006.1843|pmid=16754612|pmc=1578724}}</ref>。真核生物の起源を分子時計を用いて推測する研究も行われている<ref>{{Cite journal|last=Strassert|first=Jürgen F. H.|last2=Irisarri|first2=Iker|last3=Williams|first3=Tom A.|last4=Burki|first4=Fabien|date=2021-03-25|title=A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids|url=https://www.nature.com/articles/s41467-021-22044-z|journal=Nature Communications|volume=12|issue=1|pages=1879|language=en|doi=10.1038/s41467-021-22044-z|issn=2041-1723|pmid=33767194|pmc=7994803}}</ref><ref>{{Cite journal|last=Parfrey|first=L. W.|last2=Lahr|first2=D. J. G.|last3=Knoll|first3=A. H.|last4=Katz|first4=L. A.|date=2011-08-16|title=Estimating the timing of early eukaryotic diversification with multigene molecular clocks|url=http://www.pnas.org/cgi/doi/10.1073/pnas.1110633108|journal=Proceedings of the National Academy of Sciences|volume=108|issue=33|pages=13624–13629|language=en|doi=10.1073/pnas.1110633108|issn=0027-8424|pmid=21810989|pmc=3158185}}</ref>。ただし分子時計計算はステランなどの化学化石、および微化石の年代を基にしており、これら化石試料の選択と解釈次第で大きく計算結果が変動するため注意を要する<ref>{{Cite journal|last=Eme|first=L.|last2=Sharpe|first2=S. C.|last3=Brown|first3=M. W.|last4=Roger|first4=A. J.|date=2014-08-01|title=On the Age of Eukaryotes: Evaluating Evidence from Fossils and Molecular Clocks|url=http://cshperspectives.cshlp.org/lookup/doi/10.1101/cshperspect.a016139|journal=Cold Spring Harbor Perspectives in Biology|volume=6|issue=8|pages=a016139–a016139|language=en|doi=10.1101/cshperspect.a016139|issn=1943-0264|pmid=25085908|pmc=4107988}}</ref>(上述の否定された27億年前のステランもその例)。 |
|||
古細菌に真核生物のバイオマーカー{{Enlink|Biosignature|英語版|en}}が存在することは、古細菌起源を示唆している。[[アスガルド古細菌]]のゲノムには、真核生物の特徴である[[細胞骨格]]や複雑な細胞構造の発達に重要な役割を果たす、[[アスガルド古細菌#真核生物との関係|真核生物特有のタンパク質遺伝子]]が多く存在する。2022年、[[クライオ電子線トモグラフィー]]法によって、アスガルド古細菌が複雑な[[アクチン]]ベースの細胞骨格を持つことが明らかになり、真核生物の祖先が古細菌であることを示す最初の直接的な視覚的証拠が得られた<ref name="Rodrigues-Oliveira et al 2023">{{cite journal |vauthors=Rodrigues-Oliveira T, Wollweber F, Ponce-Toledo RI, etal. |title=Actin cytoskeleton and complex cell architecture in an Asgard archaean |journal=Nature |volume=613 |pages=332–339 |date=2023 |issue=7943 |doi=10.1038/s41586-022-05550-y|pmid=36544020 |pmc=9834061 |bibcode=2023Natur.613..332R |hdl=20.500.11850/589210 |hdl-access=free }}</ref>。 |
|||
ちなみに、[[動物]]や[[植物]]へ至る真核生物の多細胞化は真核生物自体の成立に比べて新しく、10億年前あたりを示唆する研究結果がある一方<ref name=":8" />、上述のGrypaniaが本当に多細胞性の真核生物であった場合、多細胞化の起源が大幅に遡るため、結論は出ていない。 |
|||
<!-- |
|||
== 真核生物の位置付け == |
|||
生物の分類の変遷を下に示す。真核生物の起源に関して、エオサイト説と今日提唱されている2ドメイン説は厳密には同一ではないが、古細菌と真核生物の近縁性に注目している点で同一である。また2ドメイン説においても、真核生物の名称および枠組み自体は存続している。 |
|||
古細菌から真核生物への具体的な道筋は解明されておらず、水素仮説<ref>{{Cite journal|last=Martin|first=William|last2=Müller|first2=Miklós|date=1998-03|title=The hydrogen hypothesis for the first eukaryote|url=http://www.nature.com/articles/32096|journal=Nature|volume=392|issue=6671|pages=37–41|language=en|doi=10.1038/32096|issn=0028-0836}}</ref>、リバース・フローモデル<ref>{{Cite journal|last=Spang|first=Anja|last2=Stairs|first2=Courtney W.|last3=Dombrowski|first3=Nina|last4=Eme|first4=Laura|last5=Lombard|first5=Jonathan|last6=Caceres|first6=Eva F.|last7=Greening|first7=Chris|last8=Baker|first8=Brett J.|last9=Ettema|first9=Thijs J. G.|date=2019-07|title=Proposal of the reverse flow model for the origin of the eukaryotic cell based on comparative analyses of Asgard archaeal metabolism|url=http://www.nature.com/articles/s41564-019-0406-9|journal=Nature Microbiology|volume=4|issue=7|pages=1138–1148|language=en|doi=10.1038/s41564-019-0406-9|issn=2058-5276}}</ref>、E<sup>3</sup>モデル<ref>{{Cite journal|last=Imachi|first=Hiroyuki|last2=Nobu|first2=Masaru K.|last3=Nakahara|first3=Nozomi|last4=Morono|first4=Yuki|last5=Ogawara|first5=Miyuki|last6=Takaki|first6=Yoshihiro|last7=Takano|first7=Yoshinori|last8=Uematsu|first8=Katsuyuki|last9=Ikuta|first9=Tetsuro|date=2020-01-23|title=Isolation of an archaeon at the prokaryote–eukaryote interface|url=http://www.nature.com/articles/s41586-019-1916-6|journal=Nature|volume=577|issue=7791|pages=519–525|language=en|doi=10.1038/s41586-019-1916-6|issn=0028-0836|pmid=31942073|pmc=7015854}}</ref> など多くの仮説が提唱されている<ref>{{Cite journal|last=López-García|first=Purificación|last2=Moreira|first2=David|date=2020-05|title=The Syntrophy hypothesis for the origin of eukaryotes revisited|url=http://www.nature.com/articles/s41564-020-0710-4|journal=Nature Microbiology|volume=5|issue=5|pages=655–667|language=en|doi=10.1038/s41564-020-0710-4|issn=2058-5276}}</ref>。ほとんどの仮説が、古細菌がバクテリアを取り込んだと考えているのに対して、シントロピー・モデル<ref>{{Cite journal|last=Moreira|first=David|last2=López-García|first2=Purificación|date=1998-11|title=Symbiosis Between Methanogenic Archaea and δ-Proteobacteria as the Origin of Eukaryotes: The Syntrophic Hypothesis|url=http://link.springer.com/10.1007/PL00006408|journal=Journal of Molecular Evolution|volume=47|issue=5|pages=517–530|language=en|doi=10.1007/PL00006408|issn=0022-2844}}</ref> と呼ばれる仮説のみ、バクテリア(特に[[プロテオバクテリア|デルタプロテオバクテリア]])が古細菌を取り込んだと推定しており、共生の関係性が他の説とは逆である。この説ではミトコンドリアは古細菌とは別個に取り込まれて成立したとされる。上記の説以外にも、真核生物の細胞核に類似の器官をもつ一部のバクテリア(例えば[[プランクトミケス門|プランクトミケス]])が、真核生物の起源に関与しているとする説も存在する<ref>{{Cite journal|last=Fuerst|first=John A.|last2=Sagulenko|first2=Evgeny|date=2011-06|title=Beyond the bacterium: planctomycetes challenge our concepts of microbial structure and function|url=http://www.nature.com/articles/nrmicro2578|journal=Nature Reviews Microbiology|volume=9|issue=6|pages=403–413|language=en|doi=10.1038/nrmicro2578|issn=1740-1526}}</ref>。 |
|||
コメント: エオサイト説・2分岐説は系統・起源についての説だと思うので、生物の分類の枠組みと同列に並べるのは適切でないのでは。 このような図を提示するなら少なくとももっと説明は必要。 それと「2ドメイン説」という言葉は、系統についてなのか、分類体系として2ドメインにすることまで含意するのか不明なのでは。 |
|||
=== 成立年代の推定 === |
|||
真核生物の成立年代は未確定ではあるものの、例えば真核生物に不可欠ないくつかの器官(例えば[[ミトコンドリア]]や、[[ステロール]]を含む[[細胞膜]])<ref>{{Cite journal|last=Roger|first=Andrew J.|last2=Muñoz-Gómez|first2=Sergio A.|last3=Kamikawa|first3=Ryoma|date=2017-11|title=The Origin and Diversification of Mitochondria|url=https://doi.org/10.1016/j.cub.2017.09.015|journal=Current Biology|volume=27|issue=21|pages=R1177–R1192|doi=10.1016/j.cub.2017.09.015|issn=0960-9822}}</ref><ref name=":9">{{Cite journal|last=Hoshino|first=Yosuke|last2=Gaucher|first2=Eric A.|date=2021-06-22|title=Evolution of bacterial steroid biosynthesis and its impact on eukaryogenesis|url=https://www.pnas.org/content/118/25/e2101276118|journal=Proceedings of the National Academy of Sciences|volume=118|issue=25|language=en|doi=10.1073/pnas.2101276118|issn=0027-8424|pmid=34131078|pmc=8237579}}</ref>の成立に酸素が必須なことから、真核生物は24億年前の大酸化イベント以後、[[好気呼吸|好気性]]条件下でおおまかに19億年前頃([[原生代]])には成立したとする説が有力である<ref name=":8"/>。一方で、真核生物は酸素が大気中に含まれていなかった{{Ill2|大酸化イベント|en|Great Oxidation Event}}(GOE)以前の生活スタイル([[嫌気呼吸]])も保持しており<ref>{{Cite journal|last=Müller|first=Miklós|last2=Mentel|first2=Marek|last3=van Hellemond|first3=Jaap J.|last4=Henze|first4=Katrin|last5=Woehle|first5=Christian|last6=Gould|first6=Sven B.|last7=Yu|first7=Re-Young|last8=van der Giezen|first8=Mark|last9=Tielens|first9=Aloysius G. M.|date=2012-06|title=Biochemistry and Evolution of Anaerobic Energy Metabolism in Eukaryotes|url=https://journals.asm.org/doi/10.1128/MMBR.05024-11|journal=Microbiology and Molecular Biology Reviews|volume=76|issue=2|pages=444–495|language=en|doi=10.1128/MMBR.05024-11|issn=1092-2172|pmid=22688819|pmc=3372258}}</ref><ref>{{Cite book|title=Mitochondria and Anaerobic Energy Metabolism in Eukaryotes: Biochemistry and Evolution|url=https://www.degruyter.com/document/doi/10.1515/9783110612417/html|publisher=De Gruyter|date=2020-12-07|isbn=978-3-11-061241-7|doi=10.1515/9783110612417|first=William F.|last=Martin|first2=Aloysius G. M.|last2=Tielens|first3=Marek|last3=Mentel}}</ref>、最初に誕生した真核生物は[[通性嫌気性生物]]であったと想定される。大酸化イベント以前([[太古代]])の地球にもごく少量の酸素は存在していた可能性があるが<ref>{{Cite journal|last=Catling|first=David C.|last2=Zahnle|first2=Kevin J.|date=2020-02|title=The Archean atmosphere|url=https://advances.sciencemag.org/lookup/doi/10.1126/sciadv.aax1420|journal=Science Advances|volume=6|issue=9|pages=eaax1420|language=en|doi=10.1126/sciadv.aax1420|issn=2375-2548|pmid=32133393|pmc=7043912}}</ref>、真核生物を含め好気性生物が太古代にすでに存在していたかについては、それを明確に支持する証拠は現在のところない。 |
|||
[[オーストラリア]][[頁岩]]に真核生物に特有のバイオマーカーである[[ステラン]]が含まれていることから、かつては27億年前の岩石に真核生物が存在していたことが示唆されていたが<ref name="sterane">{{cite journal |vauthors=Brocks JJ, Logan GA, Buick R, Summons RE |title=Archean molecular fossils and the early rise of eukaryotes |journal=Science |volume=285 |issue=5430 |pages=1033–1036 |date=August 1999 |pmid=10446042 |doi=10.1126/science.285.5430.1033 |bibcode=1999Sci...285.1033B |citeseerx=10.1.1.516.9123 }}</ref><ref>{{cite magazine |vauthors=Ward P |title=Mass extinctions: the microbes strike back |magazine=[[:en:New Scientist|New Scientist]] |pages=40–43 |date=9 February 2008 |url=https://www.newscientist.com/channel/life/mg19726421.900-mass-extinctions-the-microbes-strike-back.html |authorlink=Peter Ward (paleontologist) |access-date=27 August 2017 |archive-date=8 July 2008 |archive-url=https://web.archive.org/web/20080708222803/http://www.newscientist.com/channel/life/mg19726421.900-mass-extinctions-the-microbes-strike-back.html |url-status=live }}</ref><ref>{{Cite journal|last=Brocks|first=J. J.|date=1999-08-13|title=Archean Molecular Fossils and the Early Rise of Eukaryotes|url=https://www.sciencemag.org/lookup/doi/10.1126/science.285.5430.1033|journal=Science|volume=285|issue=5430|pages=1033–1036|doi=10.1126/science.285.5430.1033}}</ref><ref>{{Cite journal|last=Waldbauer|first=Jacob R.|last2=Sherman|first2=Laura S.|last3=Sumner|first3=Dawn Y.|last4=Summons|first4=Roger E.|date=2009-03|title=Late Archean molecular fossils from the Transvaal Supergroup record the antiquity of microbial diversity and aerobiosis|url=https://linkinghub.elsevier.com/retrieve/pii/S0301926808002507|journal=Precambrian Research|volume=169|issue=1-4|pages=28–47|language=en|doi=10.1016/j.precamres.2008.10.011}}</ref><ref>{{Cite journal|last=Brocks|first=Jochen J|last2=Buick|first2=Roger|last3=Summons|first3=Roger E|last4=Logan|first4=Graham A|date=2003-11|title=A reconstruction of Archean biological diversity based on molecular fossils from the 2.78 to 2.45 billion-year-old Mount Bruce Supergroup, Hamersley Basin, Western Australia|url=https://linkinghub.elsevier.com/retrieve/pii/S0016703703002096|journal=Geochimica et Cosmochimica Acta|volume=67|issue=22|pages=4321–4335|language=en|doi=10.1016/S0016-7037(03)00209-6}}</ref>、これらの太古代のバイオマーカーは後世の汚染物質であると反論されている<ref>{{Cite journal|last=French|first=Katherine L.|last2=Hallmann|first2=Christian|last3=Hope|first3=Janet M.|last4=Schoon|first4=Petra L.|last5=Zumberge|first5=J. Alex|last6=Hoshino|first6=Yosuke|last7=Peters|first7=Carl A.|last8=George|first8=Simon C.|last9=Love|first9=Gordon D.|date=2015-05-12|title=Reappraisal of hydrocarbon biomarkers in Archean rocks|url=https://www.pnas.org/content/112/19/5915|journal=Proceedings of the National Academy of Sciences|volume=112|issue=19|pages=5915–5920|doi=10.1073/pnas.1419563112|pmid=25918387|pmc=4434754}}</ref><ref>{{cite journal |vauthors=French KL, Hallmann C, Hope JM, Schoon PL, Zumberge JA, Hoshino Y, Peters CA, George SC, Love GD, Brocks JJ, Buick R, Summons RE |title=Reappraisal of hydrocarbon biomarkers in Archean rocks |journal=Proceedings of the National Academy of Sciences of the United States of America |volume=112 |issue=19 |pages=5915–5920 |date=May 2015 |pmid=25918387 |doi=10.1073/pnas.1419563112 |pmc=4434754 |bibcode=2015PNAS..112.5915F |doi-access=free }}</ref><ref>{{Cite journal|last=Rasmussen|first=Birger|last2=Fletcher|first2=Ian R.|last3=Brocks|first3=Jochen J.|last4=Kilburn|first4=Matt R.|date=2008-10|title=Reassessing the first appearance of eukaryotes and cyanobacteria|url=http://www.nature.com/articles/nature07381|journal=Nature|volume=455|issue=7216|pages=1101–1104|language=en|doi=10.1038/nature07381|issn=0028-0836}}</ref>。最も古く確かなバイオマーカーの記録は、約8億年前の[[新原生代]]のものでしかない<ref>{{cite journal |vauthors=Brocks JJ, Jarrett AJ, Sirantoine E, Hallmann C, Hoshino Y, Liyanage T |title=The rise of algae in Cryogenian oceans and the emergence of animals |journal=Nature |volume=548 |issue=7669 |pages=578–581 |date=August 2017 |pmid=28813409 |doi=10.1038/nature23457 |s2cid=205258987 |bibcode=2017Natur.548..578B }}</ref><ref>{{Cite journal|last=Hoshino|first=Yosuke|last2=Poshibaeva|first2=Aleksandra|last3=Meredith|first3=William|last4=Snape|first4=Colin|last5=Poshibaev|first5=Vladimir|last6=Versteegh|first6=Gerard J. M.|last7=Kuznetsov|first7=Nikolay|last8=Leider|first8=Arne|last9=van Maldegem|first9=Lennart|date=2017-09|title=Cryogenian evolution of stigmasteroid biosynthesis|url=https://advances.sciencemag.org/lookup/doi/10.1126/sciadv.1700887|journal=Science Advances|volume=3|issue=9|pages=e1700887|language=en|doi=10.1126/sciadv.1700887|issn=2375-2548|pmc=5606710|pmid=28948220}}</ref><ref>{{Cite journal|last=Brocks|first=Jochen J.|last2=Jarrett|first2=Amber J. M.|last3=Sirantoine|first3=Eva|last4=Hallmann|first4=Christian|last5=Hoshino|first5=Yosuke|last6=Liyanage|first6=Tharika|date=2017-08|title=The rise of algae in Cryogenian oceans and the emergence of animals|url=https://www.nature.com/articles/nature23457|journal=Nature|volume=548|issue=7669|pages=578–581|language=en|doi=10.1038/nature23457|issn=1476-4687}}</ref>。対照的に、分子時計分析によれば、ステロール生合成が23億年前にも出現したことを示唆している<ref>{{cite journal |vauthors=Gold DA, Caron A, Fournier GP, Summons RE |title=Paleoproterozoic sterol biosynthesis and the rise of oxygen |journal=Nature |volume=543 |issue=7645 |pages=420–423 |date=March 2017 |pmid=28264195 |doi=10.1038/nature21412 |hdl-access=free |s2cid=205254122 |bibcode=2017Natur.543..420G |hdl=1721.1/128450 }}</ref>。真核生物のバイオマーカーとしてのステランの性質は、一部の細菌によるステロールの産生によってさらに複雑になっている<ref>{{cite journal |vauthors=Wei JH, Yin X, Welander PV |title=Sterol Synthesis in Diverse Bacteria |journal=Frontiers in Microbiology |volume=7 |pages=990 |date=2016-06-24 |pmid=27446030 |pmc=4919349 |doi=10.3389/fmicb.2016.00990 |doi-access=free |authorlink3=Paula V. Welander }}</ref><ref>{{cite journal |vauthors=Hoshino Y, Gaucher EA |title=Evolution of bacterial steroid biosynthesis and its impact on eukaryogenesis |journal=Proceedings of the National Academy of Sciences of the United States of America |volume=118 |issue=25 |pages=e2101276118 |date=June 2021 |pmid=34131078 |pmc=8237579 |doi=10.1073/pnas.2101276118 |bibcode=2021PNAS..11801276H |doi-access=free }}</ref>。 |
|||
文案: |
|||
過去に提唱された生物の分類の枠組みと、エオサイト説・2分岐説で考えられている系統群の関係を示す。 |
|||
なお、2分岐説でも真核生物という分類群が解体されるわけではない。 |
|||
また、分類体系をどうするか、分類群の階級をどうするかといったことは系統関係とはまた別の問題がある。 |
|||
新原生代以前の真核生物の有無および実態については詳しくわかっていない。2023年、現生の真核生物がもつステロールとは化学構造がやや異なる、”より原始的な”プロトステロールが化石化したものが新原生代以前の地層に広く分布していることが発表され、これらのステロールは現生の真核生物([[クラウングループ|クラウン・グループ]])以前に存在していたステム・グループに属する生物が作り出していた可能性が指摘された<ref>{{Cite journal|last=Brocks|first=Jochen J.|last2=Nettersheim|first2=Benjamin J.|last3=Adam|first3=Pierre|last4=Schaeffer|first4=Philippe|last5=Jarrett|first5=Amber J. M.|last6=Güneli|first6=Nur|last7=Liyanage|first7=Tharika|last8=van Maldegem|first8=Lennart M.|last9=Hallmann|first9=Christian|date=2023-06-22|title=Lost world of complex life and the late rise of the eukaryotic crown|url=https://www.nature.com/articles/s41586-023-06170-w|journal=Nature|volume=618|issue=7966|pages=767–773|language=en|doi=10.1038/s41586-023-06170-w|issn=0028-0836}}</ref>。この説に従えば、現存する{{仮リンク|真核生物発生|label=真核生物の最終共通祖先|en|Eukaryogenesis}}(LECA)は新原生代まで出現しなかったことになり、それまでは真核生物の前駆段階にあたる何らかの好気性生物が長く繁栄していたことになる。一方で、プロトステロールを含めてステロール自体は細菌が究極的な起源である可能性も指摘されており<ref name=":9" />、新原生代以前のステロール(プロトステロール)を合成していた生物が何者だったのかによって、真核生物の成立過程についての理解は今後大きく変化する可能性がある。 |
|||
{| class="wikitable" style="margin-left: 0.5em; style=font-size:90%;" |
|||
!2界説 |
|||
![[界 (分類学)#三界説|3界説]] |
|||
![[5界説]] |
|||
!6界説 |
|||
!style="width:13%" |[[3ドメイン説]]<ref name="Woese_1990"> |
|||
{{cite journal|author=Woese C, Kandler O, Wheelis M|year=1990|title=Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya.(生物の自然機構について:古細菌、細菌、真核生物の3ドメインの提案)|journal=Proc Natl Acad Sci U S A|volume=87|issue=12|pages=4576-9|u=http://www.pnas.org/cgi/reprint/87/12/4576|id=PMID 2112744}}</ref> |
|||
!style="width:16%" |<!- - 2021年現在の現行説 - -> |
|||
2ドメイン説・[[エオサイト説]] |
|||
! rowspan="2" |具体例<ref>ここに載せた具体例は下記より引用:{{Cite book|ref=藤田10|author=藤田敏彦|title=動物の系統分類と進化|series=新・生命科学シリーズ|date=2010/4/28|publisher=裳華房|isbn=978-4785358426|author2=|author3=|author4=|author5=|author6=|author7=|author8=|author9=}} p91</ref> |
|||
|- |
|||
![[カール・フォン・リンネ|リンネ]]<br/>([[1735年]]) |
|||
![[エルンスト・ヘッケル|ヘッケル]]<br/>([[1866年]]) |
|||
![[ロバート・ホイタッカー|ホイタッカー]]<br/>([[1969年]]) |
|||
![[カール・ウーズ|ウーズ]]<br/>([[1977年]]) |
|||
!ウーズ<br/>([[1990年]]) |
|||
!レイクら<br/>([[1984年]]) |
|||
|- |
|||
| rowspan="3" | |
|||
| rowspan="3" style="background-color: khaki;" |'''[[原生生物]]界''' |
|||
| rowspan="2" style="background-color: lightgrey;" |'''[[モネラ界]]''' |
|||
| style="background-color: lightgrey;" |'''[[細菌|真正細菌]]界''' |
|||
| style="background-color: lightgrey;" |'''[[細菌]]''' |
|||
| style="background-color: lightgrey;" |'''[[細菌]]''' |
|||
|大腸菌、放線菌、藍色細菌 |
|||
|- |
|||
| style="background-color: #F3E0E0;" |'''[[古細菌]]界''' |
|||
| style="background-color: #F3E0E0;" |'''[[アーキア]] (古細菌)''' |
|||
| rowspan="5" style="background-color: #F3E0E0;" |'''[[アーキア]]'''<!-(古細菌)-><!- Arkarya ? -> |
|||
ただし真核生物は古細菌から分岐したもので、1行目以外が真核生物に対応 |
|||
|[[メタン菌|メタン生成菌]]、[[好熱好酸菌]] |
|||
|- |
|||
| style="background-color: khaki;" |'''[[原生生物]]界''' |
|||
| style="background-color: khaki;" |'''[[原生生物]]界''' |
|||
| rowspan="4" style="background-color: #e0d0b0;" |'''[[真核生物]]''' |
|||
|[[藻類]]、[[原生動物]]、[[変形菌|変形菌類]] |
|||
|- |
|||
| rowspan="2" style="background-color: lightgreen;" |'''[[植物]]界''' |
|||
| rowspan="2" style="background-color: lightgreen;" |'''[[植物]]界''' |
|||
| style="background-color: lightblue;" |'''[[菌類|菌界]]''' |
|||
| style="background-color: lightblue;" |'''[[菌類|菌界]]''' |
|||
|[[キノコ]]、[[カビ]]、[[地衣類|地衣植物]] |
|||
|- |
|||
| style="background-color: lightgreen;" |'''[[植物]]界''' |
|||
| style="background-color: lightgreen;" |'''[[植物]]界''' |
|||
|[[苔|コケ類]]、[[シダ植物門|シダ類]]、[[種子植物]] |
|||
|- |
|||
| style="background-color: rgb(211,211,164);" |'''[[動物]]界''' |
|||
| style="background-color: rgb(211,211,164);" |'''[[動物]]界''' |
|||
| style="background-color: rgb(211,211,164);" |'''[[動物]]界''' |
|||
| style="background-color: rgb(211,211,164);" |'''[[動物]]界''' |
|||
|[[無脊椎動物]]、[[脊椎動物]] |
|||
|} |
|||
上の表の「動物界」、「植物界」などに登場する「[[界 (分類学)|界]]」という語は、生物の分類階級の一つで、3ドメイン説が登場するまでは最上位の分類階級として位置づけられていた。 |
|||
--> |
|||
ステラン以外の真核生物の痕跡としては、真核生物由来とされる[[微化石]]が21億年前の地層から発見されている<ref>{{Cite journal|last=Albani|first=Abderrazak El|last2=Bengtson|first2=Stefan|last3=Canfield|first3=Donald E.|last4=Bekker|first4=Andrey|last5=Macchiarelli|first5=Roberto|last6=Mazurier|first6=Arnaud|last7=Hammarlund|first7=Emma U.|last8=Boulvais|first8=Philippe|last9=Dupuy|first9=Jean-Jacques|date=2010-07|title=Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago|url=https://www.nature.com/articles/nature09166|journal=Nature|volume=466|issue=7302|pages=100–104|language=en|doi=10.1038/nature09166|issn=1476-4687}}</ref>。ただし、これらの化石が真に真核生物由来かどうかはなお議論の必要がある。19億年前の地層から見つかった、コイル状の多細胞生物と推定されるGrypaniaは真核生物として一定の支持を得ている最古の化石の一つである<ref name=":8">{{Cite journal|last=Knoll|first=A.h|last2=Javaux|first2=E.j|last3=Hewitt|first3=D|last4=Cohen|first4=P|date=2006-06-29|title=Eukaryotic organisms in Proterozoic oceans|url=https://royalsocietypublishing.org/doi/10.1098/rstb.2006.1843|journal=Philosophical Transactions of the Royal Society B: Biological Sciences|volume=361|issue=1470|pages=1023–1038|doi=10.1098/rstb.2006.1843|pmid=16754612|pmc=1578724}}</ref>。真核生物の起源を分子時計を用いて推測する研究も行われている<ref>{{Cite journal|last=Strassert|first=Jürgen F. H.|last2=Irisarri|first2=Iker|last3=Williams|first3=Tom A.|last4=Burki|first4=Fabien|date=2021-03-25|title=A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids|url=https://www.nature.com/articles/s41467-021-22044-z|journal=Nature Communications|volume=12|issue=1|pages=1879|language=en|doi=10.1038/s41467-021-22044-z|issn=2041-1723|pmid=33767194|pmc=7994803}}</ref><ref>{{Cite journal|last=Parfrey|first=L. W.|last2=Lahr|first2=D. J. G.|last3=Knoll|first3=A. H.|last4=Katz|first4=L. A.|date=2011-08-16|title=Estimating the timing of early eukaryotic diversification with multigene molecular clocks|url=http://www.pnas.org/cgi/doi/10.1073/pnas.1110633108|journal=Proceedings of the National Academy of Sciences|volume=108|issue=33|pages=13624–13629|language=en|doi=10.1073/pnas.1110633108|issn=0027-8424|pmid=21810989|pmc=3158185}}</ref>。 |
|||
== 下位分類 == |
|||
[[五界説]]では、真核生物は動物、植物、菌類、原生生物の4つの[[界 (分類学)|界]]に分類されていた。<!-- 例えば、原生生物の分類体系であるLevine ''et al.'' (1980)は、国際原生動物学会の合意して図鑑や百科事典などに取り入れられていた。-->しかし近年では、分子系統解析などの研究成果を受け、真核生物の新しい分類体系が発表されている。例えば、動物と菌類は同一の系統に含まれるとして[[オピストコンタ]]にまとめられている。 |
|||
その起源が何であれ、真核生物が生態学的に優勢になったのは、ずっと後のことかもしれない。8億年前に海洋堆積物の{{Ill2|生命起源論の代替シナリオ|en|Alternative abiogenesis scenarios#Zinc world|label=亜鉛組成}}が大幅に増加したのは、原核生物に比べて[[亜鉛]]を優先的に消費し取り込む真核生物の個体数が、その起源から(遅くとも)約10億年後に増加したことに起因している<ref name="pmid29869832">{{cite journal |vauthors=Isson TT, Love GD, Dupont CL, Reinhard CT, Zumberge AJ, Asael D, Gueguen B, McCrow J, Gill BC, Owens J, Rainbird RH, Rooney AD, Zhao MY, Stueeken EE, Konhauser KO, John SG, Lyons TW, Planavsky NJ |display-authors=3 |title=Tracking the rise of eukaryotes to ecological dominance with zinc isotopes |journal=Geobiology |volume=16|issue=4|pages=341–352|date=June 2018 |pmid=29869832 |doi=10.1111/gbi.12289 |bibcode=2018Gbio...16..341I |doi-access=free }}</ref>。 |
|||
<!-- {{疑問点|date=2021年8月|title=ISOPの分類体系は 系統を重視した分類体系 であって、系統樹そのものではないでしょう。}} --> |
|||
現在提案されている分類体系として、[[国際原生生物学会]](ISOP)によるものがある。この体系は2005年に発表され<ref name=":4">[[#Adl(2005)]]</ref>2012年<ref name=":5">[[#Adl(2012)]]</ref>と2019年<ref name=":0">[[#Adl(2019)]]</ref>に2度改訂されている。以下に2019年に出版された改訂版(Adl ''et al.,'' 2019)の概観を表に示す<ref name=":0" /><ref>[[#矢﨑(2020)]]</ref>。なお真核生物の系統関係の解明は2020年現在も進展中であり<ref>{{Cite journal|last=Burki|first=Fabien|last2=Roger|first2=Andrew J.|last3=Brown|first3=Matthew W.|last4=Simpson|first4=Alastair G.B.|date=2020-01|title=The New Tree of Eukaryotes|url=https://linkinghub.elsevier.com/retrieve/pii/S0169534719302575|journal=Trends in Ecology & Evolution|volume=35|issue=1|pages=43–55|language=en|doi=10.1016/j.tree.2019.08.008}}</ref>、今後も改訂が続いていくと考えられる。 |
|||
{| class="wikitable" |
|||
| rowspan="2" |[[アモルフェア]] |
|||
{{sname||Amorphea}} |
|||
| colspan="2" |[[アメーボゾア]] |
|||
{{Sname||Amoebozoa}}(アメーバや粘菌を含む系統) |
|||
|- |
|||
| colspan="2" |[[incertae sedis]] オバゾア |
|||
{{Sname||Obazoa}}([[動物|後生動物]]や[[菌類]]を含む系統) |
|||
|- |
|||
| rowspan="3" |[[ディアフォレティケス]] |
|||
{{Sname||Diaphoretickes}} |
|||
|[[アーケプラスチダ|古色素体類]] |
|||
{{Sname||Archaeplastida}} |
|||
|[[灰色藻|灰色藻類]] 、[[紅藻|紅藻類]]、[[緑色植物亜界|緑色植物]]を含む系統 |
|||
|- |
|||
| colspan="2" |[[SARスーパーグループ|SAR]] [[:en:SAR supergroup|Sar]] |
|||
|- |
|||
| colspan="2" |[[ハプティスタ|ハプチスタ]] {{Sname||Haptista}}や[[クリプチスタ]] {{Sname||Cryptista}}などいくつかの単系統群 |
|||
|- |
|||
| colspan="3" |クルムス、[[メタモナス|メタモナダ]] 、ディスコバ、マラウィモナス類等以上に属さないいくつかの単系統群 |
|||
|}<!-- 真核生物は、アモルフェアとディアフォレティケスの2つに大別される。さらにそれらは、[[スーパーグループ (分類学)|スーパーグループ]]と呼ばれる大きな系統群を含む。スーパーグループの1つであった[[エクスカバータ]]は、 --> |
|||
* 2005年と2012年版の体系では主要な分類群の1つとして採用されていた「[[エクスカバータ]]」は、単系統性が疑わしいとされ<ref>[[#矢﨑(2020)]] p.74.</ref>、ディスコバ、メタモナダ、マラウィモナス類の3つに解体された。 |
|||
* オピストコンタ、アプソモナス類、Breviatea は単系統群を形成すると考えられ、{{sname||Obazoa}} と呼ばれる。 |
|||
== |
=== 化石 === |
||
[[細胞核]]という構造の有無が生物の分類にとって重要な差異であることは、19世紀にはすでに認識されていた。たとえば[[原生生物]]という言葉を初めて用いた[[エルンスト・ヘッケル]]は、細菌などのなんの構造も持たない生物を原生生物の中の[[モネラ]]として区別し、後に[[藍藻]]をここに含めている<ref name="Sapp2005">{{cite journal|author=Sapp, J.|year=2005|title=The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology|journal=Microbiol. Mol. Biol. Rev.|volume=69|issue=2|pages=292-305|doi=10.1128/mmbr.69.2.292-305.2005}}</ref>。しかし当時は[[動物]]と[[植物]]という差異がまず先に立っており、モネラとそれ以外という差異が注目されることはなかった。 |
|||
真核生物の起源を特定するのは困難であるが、16億3,500万年前に生息していた最古の多細胞真核生物である{{Snamei|Qingshania magnificia}}が中国北部で発見されたことは、[[クラウングループ]]の真核生物が[[古原生代]]後期([[スタテリアン|スタテリアン紀]])に起源を持つことを示唆している。約16億5,000万年前に生息していた最初期の明確な単細胞真核生物も中国北部で発見された。それらは、''Tappania plana'', ''Shuiyousphaeridium macroreticulatum'', ''Dictyosphaera macroreticulata'', ''Germinosphaera alveolata'', and ''Valeria lophostriata'' である<ref>{{Cite journal|last1=Miao |first1=L. |last2=Yin |first2=Z. |last3=Knoll |first3=A. H. |last4=Qu |first4=Y. |last5=Zhu |first5=M. |title=1.63-billion-year-old multicellular eukaryotes from the Chuanlinggou Formation in North China |year=2024 |journal=Science Advances |volume=10 |issue=4 |pages=eadk3208 |doi=10.1126/sciadv.adk3208 |doi-access=free |pmid=38266082 |pmc=10807817 }}</ref>。 |
|||
真核生物という言葉は、文献上[[エドゥアール・シャットン]]が1925年の論文で初めて用いた<ref name="Katscher2004">{{cite journal|author=Katscher, F.|year=2004|title=The History of the Terms Prokaryotes and Eukaryotes| journal=Protist|volume=155|pages=257-263|doi=10.1078/143446104774199637}}</ref>。この論文は{{snamei|Pansporella perplexa}}の分類学的位置を議論するもので、末尾の原生生物の分類表と樹形図の中で{{lang|fr|Eucaryotes}}と{{lang|fr|Procaryotes}}が示されているものの、他には何の説明もなかった<ref>{{cite journal|author=Chatton, E.|year=1925|title=Pansporella perplexa, amoebien à spores protégées parasite de daphnies|journal=Ann. Sci. Nat. Zool.|volume=8|pages=5–84|url=https://gallica.bnf.fr/ark:/12148/bpt6k55415433/f8.item}}</ref>。シャットンの弟子で後に[[ノーベル生理学・医学賞]]を受賞した[[アンドレ・ルヴォフ]]の1932年のモノグラフの冒頭には、シャットンを引用しながら原生生物を原核生物と真核生物に二分する旨の記述がある。ここでは、原核的原生生物を[[細胞核]]や[[ミトコンドリア]]がないもの、真核的原生生物を両者を持つものとしている<ref>{{cite book|author=Lwoff, A.|year=1932|title=Recherches Biochimiques sur la Nutrition des Protozoaires. Le Pouvoir de Synthèse.|publisher=Masson|locastion=Paris}}</ref>。以後、20世紀前半に[[英語]]、[[ドイツ語]]、[[フランス語]]の文献で何度か言及されてはいるが、生物を真核生物と原核生物に二分する方法は一般的な認識とは程遠かった<ref name="Katscher2004" />。たとえば{{仮リンク|ハーバート・コープランド|en|Herbert Copeland}}は1938年に細胞核がない生物を[[モネラ界]]としたが、細胞核がある生物についてはヘッケルの3界(動物界、植物界、原生生物界)をそのまま採用している<ref>{{cite journal|author=Copeland, H.|year=1938|title=The kingdoms of organisms|journal=Quart. Rev. Biol.|volume=13|issue=4|pages=383-420|doi=10.1086/394568}}</ref>。この二分法を普及させたのは、カナダ人の[[細菌学者]][[:en:Roger Yates Stanier|Roger Yates Stanier]]である。彼は1960年から翌年にかけて[[サバティカル]]で[[パスツール研究所]]に滞在し、ルヴォフとの議論の中でシャットンの二分法を知り、1962年の論文<ref>{{cite journal|author=Stanier & van Niel|year=1962|title=The concept of a bacterium|journal=Arch. Mikrobiol.|volume=42|pages=17–35|doi=10.1007/BF00425185}}</ref>で広く知らしめたのである<ref name="Sapp2005" /><ref name="Katscher2004" />。[[電子顕微鏡]]による[[微細構造]]観察が当たり前のように行われる時代になって、この二分法は広く受け入れられるようになった。 |
|||
少なくとも16.5億年前の[[アクリターク]](分類不能な微化石)も知られており、藻類の可能性がある{{Ill2|グリパニア|en|Grypania}}({{Snamei|Grypania}})の化石は21億年前のものである<ref name="Han">{{cite journal |vauthors=Han TM, Runnegar B |title=Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan |journal=Science |volume=257 |issue=5067 |pages=232–5 |date=July 1992 |pmid=1631544 |doi=10.1126/science.1631544 |bibcode=1992Sci...257..232H |url=}}</ref><ref>{{cite journal |vauthors=Knoll AH, Javaux EJ, Hewitt D, Cohen P |title=Eukaryotic organisms in Proterozoic oceans |journal=Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences |volume=361 |issue=1470 |pages=1023–1038 |date=June 2006 |pmid=16754612 |pmc=1578724 |doi=10.1098/rstb.2006.1843 }}</ref>。「[[Incertae sedis|問題の化石]]<ref name="Retallack 2013" /><!-- ここでは生物学的関係性を問題と指している (英語版 Diskagma も参照) -->」{{Snamei|[[:en:Diskagma|Diskagma]]}} は、22億年前の{{Ill2|古土壌|en|Paleosol}}から発見された<ref name="Retallack 2013">{{cite journal |vauthors=Retallack GJ, Krull ES, Thackray GD, Parkinson DH |title= Problematic urn-shaped fossils from a Paleoproterozoic (2.2 Ga) paleosol in South Africa. |journal=Precambrian Research |year=2013 |volume=235 |pages=71–87 |doi=10.1016/j.precamres.2013.05.015 |bibcode=2013PreR..235...71R }}</ref>。[[File:Diskagma butonii.jpg|thumb|約22億年前の岩石から発見された、高さ1mmに満たない地球上の化石、{{Snamei|[[:en:Diskagma|Diskagma buttonii]]}}の復元図<ref name="Retallack 2013" />]] |
|||
しかし1990年に、[[カール・ウーズ]](Carl Woese)らにより分子[[系統学|系統解析]]を用いて真核生物、細菌、古細菌の3ドメインが導入されるに伴い、真核生物・原核生物という二分法もまた過去のものとされるに至った([[:en:Three-domain_system|Three-domain system]])<ref>{{Cite journal|last=Woese|first=C R|last2=Kandler|first2=O|last3=Wheelis|first3=M L|date=1990-06|title=Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya.|url=https://pnas.org/doi/full/10.1073/pnas.87.12.4576|journal=Proceedings of the National Academy of Sciences|volume=87|issue=12|pages=4576–4579|language=en|doi=10.1073/pnas.87.12.4576|issn=0027-8424|pmc=PMC54159|pmid=2112744}}</ref>。しかしながら、真核生物以外のすべての生物の総称として、原核生物という言葉は今日でも学術論文で用いられている。一方で21世紀に入ると、真核生物は古細菌から派生して出現した系統であるという理解が普及し、生物界を細菌とそれ以外で分ける、上記とは異なる意味合いでの二分法が出現している<ref name=":10" />。 |
|||
[[ガボン]]の{{Ill2|フランスヴィルB層|en|Francevillian B Formation}}などの[[古原生代]]の{{Ill2|黒色頁岩|en|Black shale}}からは、21億年前と推定される「{{Ill2|フランスヴィル生物相|en|Francevillian biota}}」と呼ばれる「大型生物群集」を表すとされる構造物が見つかっている<ref name="El Albani Bengtson Canfield 2010">{{cite journal |vauthors=El Albani A, Bengtson S, Canfield DE, Bekker A, Macchiarelli R, Mazurier A, Hammarlund EU, Boulvais P, Dupuy JJ, Fontaine C, Fürsich FT, Gauthier-Lafaye F, Janvier P, Javaux E, Ossa FO, Pierson-Wickmann AC, Riboulleau A, Sardini P, Vachard D, Whitehouse M, Meunier A |display-authors=3 |s2cid=4331375 |title=Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago |journal=Nature |volume=466 |issue=7302 |pages=100–104 |date=July 2010 |pmid=20596019 |doi=10.1038/nature09166 |bibcode=2010Natur.466..100A }}</ref><ref name="El Albani 2023">{{cite journal |title=A search for life in Palaeoproterozoic marine sediments using Zn isotopes and geochemistry |last=El Albani |first=Abderrazak |journal=Earth and Planetary Science Letters |year=2023 |volume=623 |page=118169 |doi=10.1016/j.epsl.2023.118169|bibcode=2023E&PSL.61218169E |s2cid=258360867 |doi-access=free |url=https://hal.science/hal-04095643/file/El%20Albani%20et%20al._EPSL_2023.pdf }}</ref>。しかし、これらの構造物が化石であるかどうかについては議論があり、これらが[[偽化石]]である可能性を示唆する著者もいる<ref name="Ossa Ossa Pons Bekker Hofmann 2023">{{cite journal | last1=Ossa Ossa | first1=Frantz | last2=Pons | first2=Marie-Laure | last3=Bekker | first3=Andrey | last4=Hofmann | first4=Axel | last5=Poulton | first5=Simon W. | last6=Andersen | first6=Morten B. | last7=Agangi | first7=Andrea | last8=Gregory | first8=Daniel | last9=Reinke | first9=Christian | last10=Steinhilber | first10=Bernd | last11=Marin-Carbonne | first11=Johanna | last12=Schoenberg | first12=Ronny | display-authors=5 | title=Zinc enrichment and isotopic fractionation in a marine habitat of the c. 2.1 Ga Francevillian Group: A signature of zinc utilization by eukaryotes? | journal=Earth and Planetary Science Letters | volume=611 | date=2023 | doi=10.1016/j.epsl.2023.118147 | page=118147| doi-access=free | bibcode=2023E&PSL.61118147O | url=https://eprints.whiterose.ac.uk/197720/8/1-s2.0-S0012821X23001607-main.pdf }}</ref>。真核生物に明確に帰属される最古の化石は、中国の濮陽(ぼくよう)層群で発見された約18億年-16億年前のものである<ref>{{Cite journal |last1=Fakhraee |first1=Mojtaba |last2=Tarhan |first2=Lidya G. |last3=Reinhard |first3=Christopher T. |last4=Crowe |first4=Sean A. |last5=Lyons |first5=Timothy W. |last6=Planavsky |first6=Noah J. |date=May 2023 |title=Earth's surface oxygenation and the rise of eukaryotic life: Relationships to the Lomagundi positive carbon isotope excursion revisited |url=https://linkinghub.elsevier.com/retrieve/pii/S0012825223000879 |journal=Earth-Science Reviews |language=en |volume=240 |pages=104398 |doi=10.1016/j.earscirev.2023.104398 |bibcode=2023ESRv..24004398F |s2cid=257761993 |url-access=subscription |access-date=6 June 2023 |archive-date=2 February 2024 |archive-url=https://web.archive.org/web/20240202091425/https://www.sciencedirect.com/science/article/abs/pii/S0012825223000879?via%3Dihub |url-status=live }}</ref>。現代の生物群と明らかに関連する化石は、[[紅藻#進化・系統|紅藻類]]の形で推定12億年前に出現し始めているが、最近の研究では、[[ヴィンディヤ山脈|ヴィンディヤ盆地]]に存在する[[藻類#歴史的概観|糸状藻類]]の化石がおそらく16億年-17億年前にさかのぼるものと示唆されている<ref>{{cite journal |vauthors=Bengtson S, Belivanova V, Rasmussen B, Whitehouse M |title=The controversial "Cambrian" fossils of the Vindhyan are real but more than a billion years older |journal=Proceedings of the National Academy of Sciences of the United States of America |volume=106 |issue=19 |pages=7729–7734 |date=May 2009 |pmid=19416859 |pmc=2683128 |doi=10.1073/pnas.0812460106 |bibcode=2009PNAS..106.7729B |doi-access=free }}</ref>。 |
|||
== 系統の詳細 == |
|||
前述した[[国際原生生物学会]](ISOP)による2019年<ref name=":0" />の分類の詳細は下記のとおりである: |
|||
{{Clear}} |
|||
{| class="wikitable" |
|||
| rowspan="28" |[[アモルフェア]] |
|||
== 関連項目 == |
|||
{{sname||Amorphea}} |
|||
| rowspan="12" |[[アメーボゾア]] |
|||
* {{Ill2|真核生物のハイブリッドゲノム|en|Eukaryote hybrid genome}} |
|||
{{Sname||Amoebozoa}} |
|||
* {{Ill2|配列決定された真核生物ゲノムのリスト|en|List of sequenced eukaryotic genomes}} |
|||
| colspan="5" |[[ツブリネア綱|ツブリネア]]<ref name=":7">[[#矢嶋(2020)]] p.76.</ref> {{Sname||Tubulinea}} |
|||
* {{Ill2|パラカリオン・ミョウジネンシス|en|Parakaryon myojinensis}}(''Parakaryon myojinensis'') |
|||
|- |
|||
* [[ヴォールト (細胞小器官)]] |
|||
| rowspan="6" |エボシア<ref name=":7" />{{Sname||Evosea}} |
|||
| colspan="4" |{{Sname||Variosea}} |
|||
|- |
|||
| rowspan="3" |真正動菌類<ref>{{Cite web|和書|url=http://protist.i.hosei.ac.jp/taxonomy/Sarcodina/Mycetozoea.html|title=動菌類 Mycetozoa|accessdate=2021/08/12|publisher=法政大学}}</ref>{{Sname||Eumycetozoa}} |
|||
| colspan="3" |[[タマホコリカビ類|タマホコリカビ綱]]{{Sname||Dictyostelia}} |
|||
|- |
|||
| colspan="3" |[[変形菌]][[タマホコリカビ類|綱]] |
|||
{{Sname||Myxogastria}} |
|||
|- |
|||
| colspan="3" |{{Sname||Protosporangiida}} |
|||
|- |
|||
| colspan="4" |{{Sname||Cutosea}} |
|||
|- |
|||
| colspan="4" |[[アーケアメーバ]]{{Sname||Archamoebea}} |
|||
|- |
|||
| rowspan="5" |[[ディスコセア綱|ディスコセア]]{{Sname||Discosea}} |
|||
| colspan="4" |フラベリナ<ref>{{Cite web|和書|url=http://protist.i.hosei.ac.jp/taxonomy/Sarcodina/Lobosea/Gymnamoebia/Amoebida/Flabellina.html|title=フラベリナ亜目|accessdate=2021/08/06|publisher=[[法政大学]]原生生物情報サーバ}}</ref>{{Sname||Flabellinia}} |
|||
|- |
|||
| colspan="4" |''スティグアメーバ'' {{Sname||Stygamoebida}} |
|||
|- |
|||
| rowspan="3" |{{Sname||Centramoebia}} |
|||
| colspan="3" |[[アカントアメーバ]]{{Sname||Acanthopodida}} |
|||
|- |
|||
| colspan="3" |{{Sname||Pellitida}} |
|||
|- |
|||
| colspan="3" |{{Sname||Himatismenida}} |
|||
|- |
|||
| rowspan="16" |[[incertae sedis]] オバゾア |
|||
{{Sname||Obazoa}} |
|||
| colspan="5" |[[アプソゾア#分類|アプソモナス類]] {{sname||Apusomonadida}} |
|||
|- |
|||
| colspan="5" |[[ブレビアテア綱|ブレビアータ類]] {{Sname||Breviatea}} |
|||
|- |
|||
| rowspan="14" |[[オピストコンタ]] |
|||
{{Sname||Opisthokonta}} |
|||
| rowspan="4" |[[ホロゾア]] {{Sname||Holozoa}} |
|||
| colspan="3" |{{Sname||Ichthyosporea}} |
|||
|- |
|||
| colspan="3" |[[フィラステレア|フィラステレア綱]]{{Sname||Filasterea}} |
|||
|- |
|||
| colspan="3" |[[襟鞭毛虫]]{{Sname||Choanoflagellata}} |
|||
|- |
|||
| colspan="3" |'''[[動物|動物界]]'''{{Sname||Metazoa}} |
|||
|- |
|||
| rowspan="10" |[[Nucletmycea]] |
|||
| colspan="3" |[[ロトスファエリダ目|ラブディオフリス]]<ref>{{Cite web|和書|url=http://protist.i.hosei.ac.jp/taxonomy/Sarcodina/Heliozoea/Rotosphaerida.html|title=ラブディオフリス目|accessdate=2021/08/06|publisher=法政大学}}</ref>{{Sname||Rotosphaerida}} |
|||
|- |
|||
| rowspan="9" |'''[[菌類|菌界]]'''{{Sname||fungi}} |
|||
| colspan="2" |{{Sname||Opisthosporidia}} |
|||
|- |
|||
| colspan="2" |[[コウマクノウキン門|コウマクノウキン目]]{{Sname||Blastocladiales}} |
|||
|- |
|||
| colspan="2" |[[ツボカビ門]]{{Sname||Chytridiomycota}} |
|||
|- |
|||
| colspan="2" |[[二核菌]]亜界 ({{sname||Dikarya}}) |
|||
|- |
|||
| colspan="2" |[[サヤミドロモドキ|サヤミドロモドキ綱]]{{Sname||Monoblepharidomycetes}} |
|||
|- |
|||
| colspan="2" |[[ケカビ亜門|ケカビ門]]{{Sname||Mucoromycota}} |
|||
|- |
|||
| colspan="2" |[[ネオカリマスティクス科]]{{Sname||Neocallimastigaceae}} |
|||
|- |
|||
| colspan="2" |フクロカビ属{{Sname||Olpidium}} |
|||
|- |
|||
| colspan="2" |{{Sname||Zoopagomycota}} |
|||
|- |
|||
| rowspan="76" |[[ディアフォレティケス]] |
|||
{{Sname||Diaphoretickes}} |
|||
| rowspan="32" |[[アーケプラスチダ|古色素体類]] |
|||
{{Sname||Archaeplastida}} |
|||
| colspan="5" |[[灰色藻|灰色藻類]] {{Sname||Glaucophyta}} |
|||
|- |
|||
| rowspan="11" |[[紅藻|'''紅藻植物門''']] {{Sname||Rhodophyceae}} |
|||
| colspan="4" |[[イデユコゴメ綱|イデユコゴメ目]]{{Sname||Cyanidiales}} |
|||
|- |
|||
| rowspan="4" |原始紅藻亜門<ref name=":2">{{Cite web|和書|url=https://tonysharks.com/Seaweeds_list/Red/Rhodophyta_top.html|title=日本産海藻リスト 紅藻類|accessdate=2021/08/07}}</ref>{{Sname||Proteorhodophytina}} |
|||
| colspan="3" |[[オオイシソウ綱|オオイシソウ目]]{{Sname||Compsopogonales}} |
|||
|- |
|||
| colspan="3" |[[チノリモ綱]]{{Sname||Porphyridiophyceae}} |
|||
|- |
|||
| colspan="3" |[[ロデラ綱|ロデラ]][[チノリモ綱|綱]]{{Sname||Rhodellophyceae}} |
|||
|- |
|||
| colspan="3" |[[ベニミドロ綱|ベニミドロ目]]{{Sname||Stylonematales}} |
|||
|- |
|||
| rowspan="6" |真正紅藻亜門<ref name=":2" />{{Sname||Eurhodophytina}} |
|||
| colspan="3" |[[ウシケノリ綱|ウシケノリ目]]{{Sname||Bangiales}} |
|||
|- |
|||
| rowspan="5" |[[真正紅藻綱]]<ref name=":2" />{{Sname||Florideophycidae}} |
|||
| colspan="2" |ベニマダラ科<ref group="注">Adl et.al.2019では真正紅藻綱の直下は「亜綱」になっているが、ここだけ「{{Sname||Hildenbrandiophycidae}}」(ベニマダラ'''亜綱''')ではなく「{{Sname||Hildenbrandiaceae}}」(ベニマダラ'''科''')になっている。</ref> {{Sname||Hildenbrandiaceae}} |
|||
|- |
|||
| colspan="2" |ウミゾウメン亜綱 {{Sname||Nemaliophycidae}} |
|||
|- |
|||
| colspan="2" |サンゴモ亜綱 {{Sname||Corallinophycidae}} |
|||
|- |
|||
| colspan="2" |イタニグサ亜綱 {{Sname||Ahnfeltiophycidae}} |
|||
|- |
|||
| colspan="2" |マサゴシバリ亜綱 {{Sname||Rhodymeniophycidae}} |
|||
|- |
|||
| rowspan="20" |[[緑色植物亜界|緑色植物]] {{Sname||Chloroplastida}} |
|||
| rowspan="13" |'''[[緑藻植物門]]'''{{Sname||Chlorophyta}} |
|||
| colspan="3" |[[アオサ藻綱]]{{Sname||Ulvophycea}} |
|||
|- |
|||
| colspan="3" |[[トレボウクシア藻綱]]{{Sname||Trebouxiophyceae}} |
|||
|- |
|||
| colspan="3" |[[緑藻綱]]{{Sname||Chlorophyceae}} |
|||
|- |
|||
| colspan="3" |[[クロロデンドロン藻綱]]{{Sname||Chlorodendrophyceae}} |
|||
|- |
|||
| colspan="3" |[[ペディノ藻綱]]{{Sname||Pedinophyceae}} |
|||
|- |
|||
| colspan="3" |[[クロロピコン藻綱]]{{Sname||Chloropicophyceae}} |
|||
|- |
|||
| colspan="3" |[[ピコキスティス藻綱]]{{Sname||Picocystophyceae}} |
|||
|- |
|||
| colspan="3" |[[ピラミモナス目]]{{Sname||Pyramimonadales}} |
|||
|- |
|||
| colspan="3" |[[マミエラ藻綱]]{{Sname||Mamiellophyceae}} |
|||
|- |
|||
| colspan="3" |ネフロセルミス属{{Sname||Nephroselmis}} |
|||
|- |
|||
| colspan="3" |{{Sname||Pycnococcaceae}} |
|||
|- |
|||
| colspan="3" |[[パルモフィルム藻綱]]{{Sname||Palmophyllophyceae}} |
|||
|- |
|||
| colspan="3" |{{Sname||Streptophyta}} |
|||
|- |
|||
| rowspan="7" |[[ストレプト植物]]{{Sname||Streptophyta}} |
|||
| colspan="3" |[[クロロキブス藻綱|クロロキブス藻属]]{{Sname||Chlorokybus}} |
|||
|- |
|||
| colspan="3" |[[メソスティグマ藻綱|メソスティグマ藻]][[クロロキブス藻綱|属]]{{Sname||Mesostigma}} |
|||
|- |
|||
| colspan="3" |[[クレブソルミディウム藻綱|クレブソルミディウム藻]][[クロロキブス藻綱|綱]]{{Sname||Klebsormidiophyceae}} |
|||
|- |
|||
| rowspan="4" |フラグモプラスト植物{{Sname||Phragmoplastophyta}} |
|||
| colspan="2" |[[接合藻|ホシミドロ藻綱]]{{Sname||Zygnematophyceae}} |
|||
|- |
|||
| colspan="2" |[[コレオケーテ藻綱]]{{Sname||Coleochaetophyceae}} |
|||
|- |
|||
| colspan="2" |[[車軸藻類|シャジクモ綱]]{{Sname||Charophyceae}} |
|||
|- |
|||
| colspan="2" |'''[[陸上植物|陸上植物門]]'''{{Sname||Embryophyta}} |
|||
|- |
|||
| rowspan="35" |[[SARスーパーグループ|SAR]] <!--{{Sname||Sar}}--> |
|||
[[:en:SAR supergroup|Sar]] |
|||
| rowspan="24" |[[ストラメノパイル]] {{Sname||Stramenopiles}} |
|||
| rowspan="6" |ビギラ{{Sname||Bigyra}} |
|||
| rowspan="4" |{{Sname||Opalozoa}} |
|||
| colspan="2" |{{Sname||Nanomonadea}} |
|||
|- |
|||
| colspan="2" |オパリナ類{{Sname||Opalinata}} |
|||
|- |
|||
| colspan="2" |[[プラシディア類]]{{Sname||Placidida}} |
|||
|- |
|||
| colspan="2" |[[ビコソエカ類|ビコソエカ目]]{{Sname||Bicosoecida}} |
|||
|- |
|||
| rowspan="2" |{{Sname||Sagenista}} |
|||
| colspan="2" |[[ラビリンチュラ類|ラビリンチュラ綱]]{{Sname||Labyrinthulomycetes}} |
|||
|- |
|||
| colspan="2" |{{Sname||Pseudophyllomitidae}} |
|||
|- |
|||
| rowspan="18" |{{Sname||Gyrista}} |
|||
| colspan="3" |{{Sname||Developea}} |
|||
|- |
|||
| colspan="3" |[[サカゲツボカビ類]]{{Sname||Hyphochytriales}} |
|||
|- |
|||
| colspan="3" |[[卵菌]]{{Sname||Peronosporomycetes}} |
|||
|- |
|||
| colspan="3" |{{Sname||Pirsoniales}} |
|||
|- |
|||
| colspan="3" |アクチノフリス科{{Sname||Actinophryidae}} |
|||
|- |
|||
| rowspan="13" |[[オクロ植物|'''オクロ植物門''']]{{Sname||Ochrophyta}} |
|||
| rowspan="8" |{{Sname||Chrysista}} |
|||
|[[黄金色藻|黄金色藻綱]]{{Sname||Chrysophyceae}} |
|||
|- |
|||
|真正眼点藻綱<ref name=":3">{{Cite web|和書|url=http://www.ujssb.org/biospnum/search.php?Kingdom=Protista&Phylum=Stramenopiles&Subphylum=Heterokontophyta|title=不等毛植物類|accessdate=2021/08/07|publisher=日本分類学会連合}}</ref>{{Sname||Eustigmatales}} |
|||
|- |
|||
|[[褐藻|褐藻綱]]<ref name=":3" />{{Sname||Phaeophyceae}} |
|||
|- |
|||
|{{Sname||Phaeothamniophyceae}} |
|||
|- |
|||
|[[ラフィド藻|ラフィド藻綱]]<ref name=":3" />{{Sname||Raphidophyceae}} |
|||
|- |
|||
|{{Sname||Schizocladia}} |
|||
|- |
|||
|[[黄緑藻|黄緑藻綱]]<ref name=":3" />{{Sname||Xanthophyceae}} |
|||
|- |
|||
|{{Sname||Diatomista}} |
|||
|- |
|||
| rowspan="5" |{{Sname||Diatomista}} |
|||
|{{Sname||Bolidophyceae}} |
|||
|- |
|||
|[[珪藻]]{{Sname||Diatomeae}} |
|||
|- |
|||
|{{Sname||Dictyochophyceae}} |
|||
|- |
|||
|ペラゴ藻綱<ref name=":3" />{{Sname||Pelagophyceae}} |
|||
|- |
|||
|ピングイオ藻綱<ref name=":3" />{{Sname||Pinguiophyceae}} |
|||
|- |
|||
| rowspan="6" |[[アルベオラータ]] {{Sname||Alveolata}} |
|||
| colspan="4" |[[コルポデラ目]]{{Sname||Colpodellida}} |
|||
|- |
|||
| colspan="4" |パーキンサス目<ref>{{Cite web|和書|url=http://fishparasite.fs.a.u-tokyo.ac.jp/Perkinsus%20olseni/Perkinsus-olseni.html|title=Perkinsus olseni(パーキンサス・オルセナイ)|accessdate=2021/08/06|publisher=[[東京大学]]}}</ref>{{Sname||Perkinsida}} |
|||
|- |
|||
| colspan="4" |コルポネマ目<ref>{{Cite web|和書|url=http://protist.i.hosei.ac.jp/taxonomy/Zoomastigophora/Genus/Colponema/index.html|title=コルポネマ属 Colponema|accessdate=2021/08/06|publisher=法政大学原生生物情報サーバ}}</ref>{{Sname||Colponemida}} |
|||
|- |
|||
| colspan="4" |[[渦鞭毛藻|渦鞭毛藻上綱]]{{Sname||Dinoflagellata}} |
|||
|- |
|||
| colspan="4" |[[アピコンプレックス門]]{{Sname||Apicomplexa}} |
|||
|- |
|||
| colspan="4" |[[繊毛虫|繊毛虫門]]{{Sname||Ciliophora}} |
|||
|- |
|||
| rowspan="5" |[[リザリア]] {{Sname||Rhizaria}} |
|||
| colspan="4" |[[ケルコゾア|ケルコゾア門]]{{Sname||Cercozoa}} |
|||
|- |
|||
| colspan="4" |{{Sname||Endomyxa}}([[ネコブカビ類]]{{Sname||Phytomyxea}}を含む系統) |
|||
|- |
|||
| rowspan="2" |レタリア類<ref>{{Cite web|和書|url=http://kccn.konan-u.ac.jp/bio/syst/pages/2107b.html|title=レタリア門|accessdate=2021/08/06}}</ref>{{Sname||Retaria}} |
|||
| colspan="3" |[[有孔虫|有孔虫下門]]{{Sname||Foraminifera}} |
|||
|- |
|||
| colspan="3" |[[放散虫|放散虫門]]{{Sname||Radiolaria}} |
|||
|- |
|||
| colspan="4" |{{Sname||Tremula}} |
|||
|- |
|||
| rowspan="2" |[[ハプティスタ|ハプチスタ]] {{Sname||Haptista}} |
|||
| colspan="5" |[[ハプト藻|ハプト藻類]]{{Sname||Haptophyta}} |
|||
|- |
|||
| colspan="5" |{{Sname||Centroplasthelida}} |
|||
|- |
|||
| rowspan="2" |[[クリプチスタ]] {{Sname||Cryptista}} |
|||
| colspan="5" |{{Snamei||Palpitomonas}} |
|||
|- |
|||
| colspan="5" |[[クリプト藻|クリプト藻綱]]{{Snamei||Cryptophyceae}} |
|||
|- |
|||
| colspan="6" |[[incertae sedis]] {{Snamei||Microheliella}} |
|||
|- |
|||
| colspan="6" |[[incertae sedis]] {{Snamei||Ancoracysta}} |
|||
|- |
|||
| colspan="6" |[[incertae sedis]] [[テロネマ門|テロネマ]] {{Sname||Telonemia}} |
|||
|- |
|||
| colspan="6" |[[incertae sedis]] {{Sname|species|Rappemonads}} |
|||
|- |
|||
| colspan="6" |[[incertae sedis]] [[ピコゾア]] {{Sname||Picozoa}} |
|||
|- |
|||
| rowspan="3" |クルムス |
|||
{{Sname||CRuMs}} |
|||
| colspan="6" |コロディクティオニッズ<ref name=":6">[[#矢﨑(2020)]] p74.</ref> {{sname||Collodictyonidae}} |
|||
|- |
|||
| colspan="6" |リジフィリッズ<ref name=":6" />{{sname||Rigifilida}} |
|||
|- |
|||
| colspan="6" |マンタモナス<ref name=":6" /> {{Snamei||Mantamonadidae|Mantamonas}} |
|||
|- |
|||
| rowspan="3" |[[メタモナス|メタモナダ]] {{Sname||Metamonada}} |
|||
| colspan="6" |{{Sname||Fornicata}}([[ランブル鞭毛虫]]を含む系統) |
|||
|- |
|||
| colspan="6" |[[パラバサリア]]{{Sname||Parabasalia}} |
|||
|- |
|||
| colspan="6" |{{Sname||Preaxostyla}} |
|||
|- |
|||
| rowspan="4" |ディスコバ{{Sname||Discoba}} |
|||
| colspan="6" |[[ジャコバ類]]{{Sname||Jakobida}} |
|||
|- |
|||
| colspan="6" |[[ツクバモナス]]{{Sname||Tsukubamonadida}} |
|||
|- |
|||
| colspan="6" |[[ヘテロロボサ]]{{Sname||Heterolobosea}} |
|||
|- |
|||
| colspan="6" |[[ユーグレノゾア]]{{Sname||Discoba}} |
|||
|- |
|||
| colspan="7" |マラウィモナス {{Sname||Malawimonadidae}} |
|||
|- |
|||
| colspan="7" |アンキロモナス類 {{Sname||Ancyromonadida}} |
|||
|- |
|||
| colspan="7" |[[ヘミマスティゴフォラ]] {{Sname||Hemimastigophora}}([[スピロネマ科|スピロネマ]]を含む系統) |
|||
|} |
|||
== 脚注 == |
== 脚注 == |
||
=== 出典 === |
|||
{{脚注ヘルプ}} |
{{脚注ヘルプ}} |
||
{{Reflist|30em}} |
|||
=== 注釈 === |
|||
{{notelist2}} |
|||
=== 出典 === |
|||
{{Reflist}} |
|||
== 参考文献 == |
=== 参考文献 === |
||
{{Wikispecies|Eukaryota}} |
|||
{{Commonscat|Eukaryota}} |
|||
*[Adl(2005)] {{Cite journal|last=Adl|first=Sina M.|last2=Simpson|first2=Alastair G. B.|last3=Farmer|first3=Mark A.|last4=Andersen|first4=Robert A.|last5=Anderson|first5=O. Roger|last6=Barta|first6=John R.|last7=Bowser|first7=Samuel S.|last8=Brugerolle|first8=Guy|last9=Fensome|first9=Robert A.|date=2005-10|title=The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists|url=http://doi.wiley.com/10.1111/j.1550-7408.2005.00053.x|journal=The Journal of Eukaryotic Microbiology|volume=52|issue=5|pages=399–451|language=en|doi=10.1111/j.1550-7408.2005.00053.x|issn=1066-5234|ref=Adl(2005)}} |
*[Adl(2005)] {{Cite journal|last=Adl|first=Sina M.|last2=Simpson|first2=Alastair G. B.|last3=Farmer|first3=Mark A.|last4=Andersen|first4=Robert A.|last5=Anderson|first5=O. Roger|last6=Barta|first6=John R.|last7=Bowser|first7=Samuel S.|last8=Brugerolle|first8=Guy|last9=Fensome|first9=Robert A.|date=2005-10|title=The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists|url=http://doi.wiley.com/10.1111/j.1550-7408.2005.00053.x|journal=The Journal of Eukaryotic Microbiology|volume=52|issue=5|pages=399–451|language=en|doi=10.1111/j.1550-7408.2005.00053.x|issn=1066-5234|ref=Adl(2005)}} |
||
*[Adl(2012)] {{Cite journal|last=Adl|first=Sina M.|last2=Simpson|first2=Alastair G. B.|last3=Lane|first3=Christopher E.|last4=Lukeš|first4=Julius|last5=Bass|first5=David|last6=Bowser|first6=Samuel S.|last7=Brown|first7=Matthew W.|last8=Burki|first8=Fabien|last9=Dunthorn|first9=Micah|date=2012-09|title=The Revised Classification of Eukaryotes|url=http://doi.wiley.com/10.1111/j.1550-7408.2012.00644.x|journal=Journal of Eukaryotic Microbiology|volume=59|issue=5|pages=429–514|language=en|doi=10.1111/j.1550-7408.2012.00644.x|pmid=23020233|pmc=3483872|ref=Adl(2012)}} |
*[Adl(2012)] {{Cite journal|last=Adl|first=Sina M.|last2=Simpson|first2=Alastair G. B.|last3=Lane|first3=Christopher E.|last4=Lukeš|first4=Julius|last5=Bass|first5=David|last6=Bowser|first6=Samuel S.|last7=Brown|first7=Matthew W.|last8=Burki|first8=Fabien|last9=Dunthorn|first9=Micah|date=2012-09|title=The Revised Classification of Eukaryotes|url=http://doi.wiley.com/10.1111/j.1550-7408.2012.00644.x|journal=Journal of Eukaryotic Microbiology|volume=59|issue=5|pages=429–514|language=en|doi=10.1111/j.1550-7408.2012.00644.x|pmid=23020233|pmc=3483872|ref=Adl(2012)}} |
||
* [Adl(2019)] {{Cite journal|last=Adl|first=Sina M.|last2=Bass|first2=David|last3=Lane|first3=Christopher E.|last4=Lukeš|first4=Julius|last5=Schoch|first5=Conrad L.|last6=Smirnov|first6=Alexey|last7=Agatha|first7=Sabine|last8=Berney|first8=Cedric|last9=Brown|first9=Matthew W.|year=2019|title=Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes|url=https://onlinelibrary.wiley.com/doi/abs/10.1111/jeu.12691|journal=Journal of Eukaryotic Microbiology|volume=66|language=en|doi=10.1111/jeu.12691|issn=1066-5234|pmid=30257078|pmc=6492006|ref=Adl(2019)}} |
* [Adl(2019)] {{Cite journal|last=Adl|first=Sina M.|last2=Bass|first2=David|last3=Lane|first3=Christopher E.|last4=Lukeš|first4=Julius|last5=Schoch|first5=Conrad L.|last6=Smirnov|first6=Alexey|last7=Agatha|first7=Sabine|last8=Berney|first8=Cedric|last9=Brown|first9=Matthew W.|year=2019|title=Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes|url=https://onlinelibrary.wiley.com/doi/abs/10.1111/jeu.12691|journal=Journal of Eukaryotic Microbiology|volume=66|language=en|doi=10.1111/jeu.12691|issn=1066-5234|pmid=30257078|pmc=6492006|ref=Adl(2019)}} |
||
* [矢﨑(2020)] {{Citation|title=真核生物の高次分類体系の改訂―Adl et al. (2019)について―|url=https://doi.org/10.19004/taxa.48.0_71|publisher=日本動物分類学会|date=2020-02-29|accessdate=2020-03-24|doi=10.19004/taxa.48.0_71|language=ja|first=裕規|last=矢﨑|first2=智之|last2=島野|ref=矢﨑(2020)}} |
* [矢﨑(2020)] {{Citation|title=真核生物の高次分類体系の改訂―Adl et al. (2019)について―|url=https://doi.org/10.19004/taxa.48.0_71|publisher=日本動物分類学会|date=2020-02-29|accessdate=2020-03-24|doi=10.19004/taxa.48.0_71|language=ja|first=裕規|last=矢﨑|first2=智之|last2=島野|ref=矢﨑(2020)}} |
||
== 外部リンク == |
|||
{{Wikispecies|Eukaryota}} |
|||
{{Commonscat|Eukaryota}} |
|||
* {{EOL}} |
|||
{{自然}} |
{{自然}} |
||
{{Normdaten}} |
{{Normdaten}} |
||
{{Taxonbar|from=Q19088}} |
{{Taxonbar|from=Q19088}} |
||
<!-- |
|||
{{Eukaryota}} |
|||
{{Life on Earth}} |
|||
{{Organisms et al.}} |
|||
--> |
|||
{{デフォルトソート:しんかくせいふつ}} |
{{デフォルトソート:しんかくせいふつ}} |
||
| 483行目: | 340行目: | ||
[[Category:真核生物|*]] |
[[Category:真核生物|*]] |
||
[[Category:生物学]] |
[[Category:生物学]] |
||
[[Category:ドメイン (分類学)]] |
|||
[[Category:生物学用語]] |
|||
2024年4月5日 (金) 13:04時点における版
| 真核生物 Eukaryota | |||
|---|---|---|---|
| 分類 | |||
| |||
| 学名 | |||
| Eukaryota | |||
| シノニム | |||
| 和名 | |||
| 真核生物 (しんかくせいぶつ) | |||
| 英名 | |||
| Eukaryote | |||
| スーパーグループと界[4] | |||






.jpg)
.jpg)
真核生物(しんかくせいぶつ、羅: Eukaryota、英: eukaryotes)は、生物学のドメイン Eukaryota あるいは Eukarya を構成し、細胞の中に核と呼ばれる細胞小器官を持つ生物である。すべての動物、植物、菌類、そして多くの単細胞生物は真核生物である。真核生物は、原核生物の2つのグループすなわち細菌と古細菌と並び、生命体の主要なグループを構成している。真核生物は生物の個体数としては少数派であるが、一般的にはるかに大きいので、その集団的な地球規模でのバイオマスは原核生物よりもはるかに大きい。
真核生物は表面上、アスガルド古細菌に属する古細菌が進化して出現したように見える。このことは、生命のドメインは細菌と古細菌の2つだけで、真核生物は古細菌の中に組み込まれていることを意味する。真核生物が最初に出現したのは古原生代のことで、おそらくは鞭毛のある細胞と考えられる。有力な進化理論によれば、真核生物は、嫌気性のアスガルド古細菌と好気性のシュードモナス門(旧: プロテオバクテリア)の共生によって誕生し、後者からミトコンドリアが形成されたという。シアノバクテリアとの共生による2回目のエピソードで、葉緑体を持つ植物の祖先が誕生した。
真核細胞(真核生物の細胞)は、核、小胞体、ゴルジ装置などの膜結合細胞小器官を持つ。真核生物には単細胞と多細胞とがある。それに比べ、原核生物は一般的に単細胞である。単細胞の真核生物は原生生物と呼ばれることもある。真核生物は有糸分裂による無性生殖と、減数分裂と配偶子融合(受精)による有性生殖の両方を行うことができる。
多様性
真核生物は、直径3マイクロメートルに満たないピコゾア門のような微小な単一細胞から[5]、体重190トン、体長33.6メートル(110 ft)のシロナガスクジラのような動物[6]、あるいは高さ120メートルにもなる(390 ft)のセコイアのような植物まで、形態的に多様性に富むさまざまな生物を指す[7]。多くの真核生物は単細胞生物で、原生生物と呼ばれる非公式なグループにはこれらの多くが含まれるが、ジャイアントケルプ(Macrocystis)のような長さ61メートル(200 ft)にもなる多細胞生物もいる[8]。多細胞の真核生物には、動物、植物、真菌類が含まれるが、やはりこれらのグループにも多くの単細胞種が含まれる[9]。真核生物の細胞は通常、原核生物(細菌や古細菌)よりもはるかに大きく、その体積は約10,000倍である[10][11]。真核生物は生物の数の中では少数派にすぎないが、その多くがはるかに大きいため、それらの世界全体のバイオマス(468ギガトン)は、原核生物(77ギガトン)よりもはるかに大きく、植物だけで地球の総バイオマスの81%以上を占めている[12]。
- 真核生物の大きさは、単細胞から何トンもある生物までさまざまである
-

-

-


.jpeg)
.jpg)
真核生物は多彩な系統であり、主に微細な生物から構成されている[13]。多細胞性は何らかの形で、真核生物の中で少なくとも25回は独立して進化してきた[14][15]。複雑な多細胞生物は、アメーバ属の集合体である粘菌類を除けば、動物、真菌類、褐藻類、紅藻類、緑藻類、陸上植物の6つの真核生物の系統の中で進化してきたにすぎない[16]。真核生物はゲノムの類似性に基づいてグループ分けされているため、グループには目に見える共通の特徴がないことが多い[13]。
特徴
核
真核生物の決定的な特徴は、その細胞が核を持っていることである。真核(eukaryote)という用語は、ギリシャ語のεὖ(eu、よく、うまく)とκάρυον(karyon、仁、核)からその名前がつけられた[17]。真核細胞には、細胞小器官と呼ばれるさまざまな内膜結合構造と、細胞の組織と形状を規定する細胞骨格がある。核は細胞のDNAを保持しており、染色体と呼ばれる線状の束に分かれている[18]。これらの染色体は、真核生物に特有の有糸分裂の過程で核分裂が起こる際、微小管紡錘体によって2つの同じ集まりに分離される[19]。
生化学
真核生物は多くの点で原核生物とは異なっており、たとえば、ステラン合成のような独特な生化学的経路を持っている[20]。真核生物のそのシグネチャータンパク質は、他の生命ドメインのタンパク質とは相同性をもたないが、真核生物の間では普遍的なもののようである。これらのタンパク質には、細胞骨格、複雑な転写機構、膜選別システム、核膜孔、および生化学的経路におけるいくつかの酵素などである[21]。
内部膜


真核生物の細胞にはさまざまな膜結合構造があり、それらの集まりが内膜系を形成している[22]。小胞や液胞と呼ばれる単純な区画は、他の膜からの出芽によって形成される。多くの細胞は、エンドサイトーシスという過程(外膜が陥入してからつまみ取るように小胞を形成する)を通じて食物やその他の物質を摂取する[23]。それに対して、エキソサイトーシスによって小胞から放出される細胞産物もある[24]。
核は核膜と呼ばれる二重膜に囲まれており、核膜孔が物質の出入りを可能にしている[25]。核膜のさまざまな管状や板状の延長部分が小胞体を形成し、タンパク質の輸送と成熟に関与している。粗面小胞体は、タンパク質を合成するリボソームで覆われた小胞体である。生成したタンパク質は内部空間あるいは内腔に入り、その後一般に、滑面小胞体から出芽した小胞に取り込まれる[26]。ほとんどの真核生物では、これらのタンパク質を輸送する小胞は放出され、ゴルジ装置と呼ばれる扁平槽(シスターネが積み重なった小器官でさらに修飾される[27]。
小胞は特殊化することもあり、たとえばリソソームは、細胞質内の生体分子を分解する消化酵素を含んでいる[28]。
ミトコンドリア

ミトコンドリアは真核細胞に存在する細胞小器官である。ミトコンドリアは、通称「細胞の発電所[29]」と呼ばれ、糖や脂肪を酸化してエネルギーを貯蔵するアデノシン三リン酸(ATP)分子を生成し、エネルギーを供給する機能を持つ[30][31]。ミトコンドリアは、リン脂質二重膜でできた2つの膜で覆われ、内側の膜はクリステという折り畳まれた構造になっていて、そこで好気呼吸が行われる[32]。
ミトコンドリアは独自のDNAを持ち、そのDNAは起源とする細菌DNAと構造的に類似しており、真核生物のRNAよりも細菌のRNAに近い構造的のRNAを生成するrRNAとtRNAの遺伝子をコードしている[33]。
一部の真核生物、たとえばメタモナス類のジアルジア属(Giardia)やトリコモナス(Trichomonas)、アメーバ動物門のペロミクサ(Pelomyxa)はミトコンドリアを欠いているように見えるが、いずれもハイドロジェノソームやマイトソームのようなミトコンドリア由来の細胞小器官を持っており、ミトコンドリアは二次的に失われたものである[34]。これらは細胞質内の酵素作用によってエネルギーを得ている[35][34]。
プラスチド

植物やさまざまな藻類は、ミトコンドリアだけでなくプラスチドも持っている。プラスチドは、ミトコンドリアと同様に独自のDNAを持ち、内部共生(この場合はシアノバクテリア)から進化した。それらは通常、葉緑体の形を取り、シアノバクテリアのようにクロロフィルを含み、光合成によってグルコースなどの有機化合物を生成する。また、栄養素の貯蔵を担うものもある。プラスチドはおそらく単一の起源を持つが、すべてのプラスチドを持つグループが密接に関連しているわけではない。それどころか、真核生物の中には、二次的な内部共生あるいは摂取によって、他の生物からそれらを獲得したものもある[36]。光合成細胞や葉緑体の捕獲と隔離、すなわち盗葉緑体化は、現代の多くの種類の真核生物で見られる[37][38]。
細胞骨格

細胞骨格は、細胞が動いたり、形を変化させたり、物質の輸送を可能にするモーター構造のために、剛構造と結合点を提供する。モーター構造はアクチンのマイクロフィラメント(微小線維)であり、α-アクチニン、フィンブリン、フィラミンなどのアクチン結合タンパク質が膜下の皮質や繊維束に存在する。微小管のモータータンパク質、ダイニンとキネシン、そしてアクチンフィラメントのミオシンが、ネットワークに動的な特性を与える[39][40]。
多くの真核生物は、鞭毛と呼ばれる細長い運動性の細胞質突起、あるいは繊毛と呼ばれる多数の短い構造を持っている。これらの細胞小器官は、運動、摂食、感覚などさまざまに関与している。それらは主にチューブリンから構成され、原核生物の鞭毛とはまったく異なる。これらは中心小体から生成する微小管の束によって支えられており、2本の1本鎖を9本の2本鎖が取り囲むように配列しているのが特徴である。鞭毛は、ストラメノパイル(Stramenopiles)の多くに見られるように、管状小毛(マスチゴネマ)を持つこともある。それらの内部は細胞質と連続している[41][42]。
中心小体は、鞭毛を持たない細胞や細胞群でもよく存在するが、針葉樹類や顕花植物にはどちらもない。これらは一般に、さまざまな微小管性鞭毛根を生じさせるグループに存在する。これらは細胞骨格の主要な構成要素を形成し、しばしば数回の細胞分裂の過程で組み立てられ、一方の鞭毛は親から受け継ぎ、もう一方はそこから派生する。中心小体は核分裂の際に紡錘体の形成に関与する[43]。
細胞壁
植物、藻類、真菌類、そしてほとんどのクロムアルベオラータ類の細胞は細胞壁に囲まれているが、動物の細胞は細胞壁に囲まれていない。これは細胞膜の外側にある層で、細胞を構造的に支え、保護し、濾過機構を提供する。細胞壁はまた、水が細胞内に侵入したときの過膨張を防ぐ役割も果たす[44]。
陸上植物の一次細胞壁を構成する主な多糖類は、セルロース、ヘミセルロース、ペクチンである。セルロース・ミクロフィブリルはヘミセルロースと結合し、ペクチン・マトリックスに埋め込まれている。一次細胞壁で最も一般的なヘミセルロースはキシログルカンである[45]。
有性生殖

真核生物は有性生殖を伴う生活環を持ち、各細胞に染色体が1つずつしか存在しない単相と、各細胞に染色体が2つずつ存在する複相とを交互に繰り返す。複相は、卵子と精子などの2つの配偶子が融合して、接合子を形成することで成立する。この接合子は、有糸分裂によって細胞分裂を繰り返しながら成体に成長し、ある段階で染色体数を減らして遺伝的変動を生み出す減数分裂によって単数体配偶子を形成する[46]。この様式にはかなりの多様性がある。植物には単数体と二倍体の両方の多細胞期がある[47]。真核生物は原核生物よりも代謝率が低く、世代時間が長くなるが、これは真核生物が原核生物よりもはるかに大きく、体積に対する表面積の比が小さいからである[48]。
有性生殖の進化は、真核生物の原初的な特徴という可能性がある。系統学的分析に基づき、DacksとRogerは通性性(facultative sex)がこのグループの共通祖先に存在したと提唱している[49]。膣トリコモナス(Trichomonas vaginalis)と腸鞭毛虫(Giardia intestinalis、あるいはランブル鞭毛虫(Giardia duodenalis))という、以前は無性であると考えられていた2つの生物には、減数分裂で機能するコア遺伝子セットが存在する[50][51]。この2種は、真核生物の進化系統樹の初期に分岐した系統の子孫であることから、コア減数分裂遺伝子、ひいては性が真核生物の共通祖先に存在した可能性がある[50][51]。リーシュマニア(Leishmania)寄生虫など、かつては無性であると考えられていた種にも性周期がある[52]。以前は無性生物と考えられていたアメーバは、古代は有性生物であり、現在の無性群体は最近進化した可能性が高い[53]。
進化
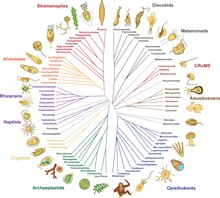
分類の歴史
古代、アリストテレスやテオプラストスは、動物と植物という2つの生物の系統を識別していた。これらの系統は、18世紀にリンネによって界(Kingdom)という分類学的な階級が与えられた。リンネは、真菌類を植物に含めることに若干の条件をつけたが、後に、真菌類はまったく別個のもので、独立した界を持つに値することがわかった[55]。さまざまな単細胞の真核生物が知られるようになった当初、それらは植物や動物と一緒にされていた。1818年、ドイツの生物学者ゲオルク・A・ゴルトフスは、繊毛虫のような生物を指すために原生動物(Protozoa)という言葉を作り[56]、このグループは、1866年にエルンスト・ヘッケルがすべての単細胞真核生物を包括する界、原生生物界(Protista)を作るまで拡張された[57][58][59]。こうして真核生物は4つの界に分類された。
当時、原生生物は「原始的な形態」であり、原始的な単細胞の性質が合併した進化の一段階であると考えられていた[58]。
19世紀にはすでに、核という構造の有無が生物の分類にとって重要な差異であることは認識されていた。エルンスト・ヘッケルは、細菌などのなんの構造も持たない生物を原生生物の中のモネラとして区別し、後にシアノバクテリアをここに含めている[60]。しかし当時は動物と植物という差異がまず先に立っており、モネラとそれ以外という差異が注目されることはなかった。
真核生物という言葉は、文献上エドゥアール・シャットンが1925年の論文で初めて用いた[61]。この論文はPansporella perplexaの分類学的位置を議論するもので、末尾の原生生物の分類表と樹形図の中でEucaryotesとProcaryotesが示されているものの、他には何の説明もなかった[62]。シャットンの弟子で後にノーベル生理学・医学賞を受賞したアンドレ・ルヴォフの1932年のモノグラフの冒頭には、シャットンを引用しながら原生生物を原核生物と真核生物に二分する旨の記述がある。ここでは、原核的原生生物を細胞核やミトコンドリアがないもの、真核的原生生物を両者を持つものとしている[63]。以後、20世紀前半に英語、ドイツ語、フランス語の文献で何度か言及されてはいるが、生物を真核生物と原核生物に二分する方法は一般的な認識とは程遠かった[61]。たとえばハーバート・コープランドは1938年に細胞核がない生物をモネラ界としたが、細胞核がある生物についてはヘッケルの3界(動物界、植物界、原生生物界)をそのまま採用している[64]。この二分法を普及させたのは、カナダ人の細菌学者 ロジャー・スタニエである。彼は1960年から翌年にかけてサバティカルでパスツール研究所に滞在し、ルヴォフとの議論の中でシャットンの二分法を知り、1962年の論文[65]で広く知らしめたのである[60][61]。電子顕微鏡による微細構造観察が当たり前のように行われる時代になって、この二分法は広く受け入れられるようになった。
生命の樹(系統樹も参照)における最古の分岐の理解は、DNAの塩基配列の決定によって初めて実質的に進展し、1990年にカール・ウーズ、オットー・カントラー、マーク・ウィーリスらが提唱した最上位の階級を(界ではなく)ドメインとする体系(3ドメイン系)が導かれた。彼らは、すべての真核生物の界を「Eucarya」ドメインに統合したが、「真核生物は今後も受け入れられる一般的な同義語である」と述べている[1][66]。1996年、進化生物学者のリン・マーギュリスは、界とドメインを「包括的」な名前に置き換えて「共生に基づく系統発生」を作ることを提案し、「真核生物(共生由来の有核生物)」という説明を加えた[2]。
しかしながら、真核生物以外のすべての生物の総称として、原核生物という言葉は今日でも学術論文で用いられている。一方で21世紀に入ると、真核生物は古細菌から派生して出現した系統であるという理解が普及し、生物界を細菌とそれ以外で分ける、上記とは異なる意味合いでの二分法が出現している[67]。
系統発生
2014年までに、過去20年間の系統学的研究から、大まかな合意が生まれはじめた[9][68]。真核生物の大部分は、アモルフェア(Amorphea、ユニコント仮説に似た構成)と、植物とほとんどの藻類系統が含まれるDiphoda(旧: バイコンタ)と呼ばれる2つの大きなクレードのいずれかに分類される。第3の主要グループであるエクスカバータ(Excavata)は、側系統であるため、正式な群としては放棄された[4]。以下の提案された系統発生図には、エクスカバータの1つの群(ディスコバ、Discoba)のみが含まれ[69]、ピコゾア門(Picozoa)は紅藻(Rhodophyta)の近縁種であるという2021年の提案が取り入れられている[70]。プロヴォラ(Provora)は2022年に発見された微生物捕食者の群である[3]。
| 真核生物/Eukaryotes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2200 mya |
この系統発生図は上界とそのステムグループの一つの見方を示す[69][71][72][13]。メタモナーダは位置づけが困難で、ディスコバあるいはマラウィモナスの姉妹という可能性がある[13]。
真核生物の起源

すべての複雑な細胞とほぼすべての多細胞生物が真核生物に含まれることから、真核生物の起源、すなわち真核発生(eukaryogenesis)は、生命の進化における画期的な出来事であった。真核生物の最終共通祖先(LECA)とは、現生するすべての真核生物の起源と仮定されるもので[74]、単一の個体ではなく生物学的な集団であった可能性が高い[75]。LECAは、核に加え、少なくとも1つの中心小体と鞭毛、通性好気性ミトコンドリア、性(減数分裂と異型配偶子融合)、キチンまたはセルロースの細胞壁を持つ休眠嚢胞、そしてペルオキシソームを持つ原生生物であったと考えられている[76][77][78]。
運動性の嫌気性古細菌と好気性アルファプロテオバクテリア綱の内共生併合によって、ミトコンドリア持つLECAそしてすべての真核生物が誕生した。さらにその後、シアノバクテリアとの2回目の内共生により、葉緑体を持つ植物の祖先が誕生した[73]。
古細菌に真核生物のバイオマーカー (en:英語版) が存在することは、古細菌起源を示唆している。アスガルド古細菌のゲノムには、真核生物の特徴である細胞骨格や複雑な細胞構造の発達に重要な役割を果たす、真核生物特有のタンパク質遺伝子が多く存在する。2022年、クライオ電子線トモグラフィー法によって、アスガルド古細菌が複雑なアクチンベースの細胞骨格を持つことが明らかになり、真核生物の祖先が古細菌であることを示す最初の直接的な視覚的証拠が得られた[79]。
古細菌から真核生物への具体的な道筋は解明されておらず、水素仮説[80]、リバース・フローモデル[81]、E3モデル[82] など多くの仮説が提唱されている[83]。ほとんどの仮説が、古細菌がバクテリアを取り込んだと考えているのに対して、シントロピー・モデル[84] と呼ばれる仮説のみ、バクテリア(特にデルタプロテオバクテリア)が古細菌を取り込んだと推定しており、共生の関係性が他の説とは逆である。この説ではミトコンドリアは古細菌とは別個に取り込まれて成立したとされる。上記の説以外にも、真核生物の細胞核に類似の器官をもつ一部のバクテリア(例えばプランクトミケス)が、真核生物の起源に関与しているとする説も存在する[85]。
成立年代の推定
真核生物の成立年代は未確定ではあるものの、例えば真核生物に不可欠ないくつかの器官(例えばミトコンドリアや、ステロールを含む細胞膜)[86][87]の成立に酸素が必須なことから、真核生物は24億年前の大酸化イベント以後、好気性条件下でおおまかに19億年前頃(原生代)には成立したとする説が有力である[88]。一方で、真核生物は酸素が大気中に含まれていなかった大酸化イベント(GOE)以前の生活スタイル(嫌気呼吸)も保持しており[89][90]、最初に誕生した真核生物は通性嫌気性生物であったと想定される。大酸化イベント以前(太古代)の地球にもごく少量の酸素は存在していた可能性があるが[91]、真核生物を含め好気性生物が太古代にすでに存在していたかについては、それを明確に支持する証拠は現在のところない。
オーストラリア頁岩に真核生物に特有のバイオマーカーであるステランが含まれていることから、かつては27億年前の岩石に真核生物が存在していたことが示唆されていたが[20][92][93][94][95]、これらの太古代のバイオマーカーは後世の汚染物質であると反論されている[96][97][98]。最も古く確かなバイオマーカーの記録は、約8億年前の新原生代のものでしかない[99][100][101]。対照的に、分子時計分析によれば、ステロール生合成が23億年前にも出現したことを示唆している[102]。真核生物のバイオマーカーとしてのステランの性質は、一部の細菌によるステロールの産生によってさらに複雑になっている[103][104]。
新原生代以前の真核生物の有無および実態については詳しくわかっていない。2023年、現生の真核生物がもつステロールとは化学構造がやや異なる、”より原始的な”プロトステロールが化石化したものが新原生代以前の地層に広く分布していることが発表され、これらのステロールは現生の真核生物(クラウン・グループ)以前に存在していたステム・グループに属する生物が作り出していた可能性が指摘された[105]。この説に従えば、現存する真核生物の最終共通祖先(LECA)は新原生代まで出現しなかったことになり、それまでは真核生物の前駆段階にあたる何らかの好気性生物が長く繁栄していたことになる。一方で、プロトステロールを含めてステロール自体は細菌が究極的な起源である可能性も指摘されており[87]、新原生代以前のステロール(プロトステロール)を合成していた生物が何者だったのかによって、真核生物の成立過程についての理解は今後大きく変化する可能性がある。
ステラン以外の真核生物の痕跡としては、真核生物由来とされる微化石が21億年前の地層から発見されている[106]。ただし、これらの化石が真に真核生物由来かどうかはなお議論の必要がある。19億年前の地層から見つかった、コイル状の多細胞生物と推定されるGrypaniaは真核生物として一定の支持を得ている最古の化石の一つである[88]。真核生物の起源を分子時計を用いて推測する研究も行われている[107][108]。
その起源が何であれ、真核生物が生態学的に優勢になったのは、ずっと後のことかもしれない。8億年前に海洋堆積物の亜鉛組成が大幅に増加したのは、原核生物に比べて亜鉛を優先的に消費し取り込む真核生物の個体数が、その起源から(遅くとも)約10億年後に増加したことに起因している[109]。
化石
真核生物の起源を特定するのは困難であるが、16億3,500万年前に生息していた最古の多細胞真核生物であるQingshania magnificiaが中国北部で発見されたことは、クラウングループの真核生物が古原生代後期(スタテリアン紀)に起源を持つことを示唆している。約16億5,000万年前に生息していた最初期の明確な単細胞真核生物も中国北部で発見された。それらは、Tappania plana, Shuiyousphaeridium macroreticulatum, Dictyosphaera macroreticulata, Germinosphaera alveolata, and Valeria lophostriata である[110]。
少なくとも16.5億年前のアクリターク(分類不能な微化石)も知られており、藻類の可能性があるグリパニア(Grypania)の化石は21億年前のものである[111][112]。「問題の化石[113]」Diskagma は、22億年前の古土壌から発見された[113]。

ガボンのフランスヴィルB層などの古原生代の黒色頁岩からは、21億年前と推定される「フランスヴィル生物相」と呼ばれる「大型生物群集」を表すとされる構造物が見つかっている[114][115]。しかし、これらの構造物が化石であるかどうかについては議論があり、これらが偽化石である可能性を示唆する著者もいる[116]。真核生物に明確に帰属される最古の化石は、中国の濮陽(ぼくよう)層群で発見された約18億年-16億年前のものである[117]。現代の生物群と明らかに関連する化石は、紅藻類の形で推定12億年前に出現し始めているが、最近の研究では、ヴィンディヤ盆地に存在する糸状藻類の化石がおそらく16億年-17億年前にさかのぼるものと示唆されている[118]。
関連項目
- 真核生物のハイブリッドゲノム
- 配列決定された真核生物ゲノムのリスト
- パラカリオン・ミョウジネンシス(Parakaryon myojinensis)
- ヴォールト (細胞小器官)
脚注
出典
- ^ a b “Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya”. Proceedings of the National Academy of Sciences of the United States of America 87 (12): 4576–4579. (June 1990). Bibcode: 1990PNAS...87.4576W. doi:10.1073/pnas.87.12.4576. PMC 54159. PMID 2112744.
- ^ a b Margulis, Lynn (6 February 1996). “Archaeal-eubacterial mergers in the origin of Eukarya: phylogenetic classification of life”. Proceedings of the National Academy of Sciences 93 (3): 1071–1076. Bibcode: 1996PNAS...93.1071M. doi:10.1073/pnas.93.3.1071. PMC 40032. PMID 8577716.
- ^ a b “Microbial predators form a new supergroup of eukaryotes”. Nature 612 (7941): 714–719. (December 2022). Bibcode: 2022Natur.612..714T. doi:10.1038/s41586-022-05511-5. PMID 36477531.
- ^ a b “Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes”. The Journal of Eukaryotic Microbiology 66 (1): 4–119. (January 2019). doi:10.1111/jeu.12691. PMC 6492006. PMID 30257078.
- ^ Seenivasan, Ramkumar; Sausen, Nicole; Medlin, Linda K.; Melkonian, Michael (26 March 2013). “Picomonas judraskeda Gen. Et Sp. Nov.: The First Identified Member of the Picozoa Phylum Nov., a Widespread Group of Picoeukaryotes, Formerly Known as 'Picobiliphytes'”. PLOS ONE 8 (3): e59565. Bibcode: 2013PLoSO...859565S. doi:10.1371/journal.pone.0059565. PMC 3608682. PMID 23555709.
- ^ Wood, Gerald (1983). The Guinness Book of Animal Facts and Feats. Enfield, Middlesex : Guinness Superlatives. ISBN 978-0-85112-235-9
- ^ Earle CJ: “Sequoia sempervirens”. The Gymnosperm Database (2017年). 2016年4月1日時点のオリジナルよりアーカイブ。2017年9月15日閲覧。
- ^ van den Hoek, C.; Mann, D.G.; Jahns, H.M. (1995). Algae An Introduction to Phycology. Cambridge: Cambridge University Press. ISBN 0-521-30419-9. オリジナルの10 February 2023時点におけるアーカイブ。 2023年4月7日閲覧。
- ^ a b “The eukaryotic tree of life from a global phylogenomic perspective”. Cold Spring Harbor Perspectives in Biology 6 (5): a016147. (May 2014). doi:10.1101/cshperspect.a016147. PMC 3996474. PMID 24789819.
- ^ DeRennaux, B. (2001). “Eukaryotes, Origin of”. Encyclopedia of Biodiversity. 2. Elsevier. pp. 329–332. doi:10.1016/b978-0-12-384719-5.00174-x. ISBN 9780123847201
- ^ “Deep-sea microorganisms and the origin of the eukaryotic cell”. Japanese Journal of Protozoology 47 (1,2): 29–48. (2014). オリジナルの9 August 2017時点におけるアーカイブ。.
- ^ Bar-On, Yinon M.; Phillips, Rob; Milo, Ron (2018-05-17). “The biomass distribution on Earth”. Proceedings of the National Academy of Sciences 115 (25): 6506–6511. Bibcode: 2018PNAS..115.6506B. doi:10.1073/pnas.1711842115. ISSN 0027-8424. PMC 6016768. PMID 29784790.
- ^ a b c d Burki, Fabien; Roger, Andrew J.; Brown, Matthew W.; Simpson, Alastair G.B. (2020). “The New Tree of Eukaryotes”. Trends in Ecology & Evolution (Elsevier BV) 35 (1): 43–55. doi:10.1016/j.tree.2019.08.008. ISSN 0169-5347. PMID 31606140.
- ^ Grosberg, RK; Strathmann, RR (2007). “The evolution of multicellularity: A minor major transition?”. Annu Rev Ecol Evol Syst 38: 621–654. doi:10.1146/annurev.ecolsys.36.102403.114735. オリジナルの14 March 2023時点におけるアーカイブ。 2023年4月8日閲覧。.
- ^ Parfrey, L.W.; Lahr, D.J.G. (2013). “Multicellularity arose several times in the evolution of eukaryotes”. BioEssays 35 (4): 339–347. doi:10.1002/bies.201200143. PMID 23315654. オリジナルの25 July 2014時点におけるアーカイブ。 2023年4月8日閲覧。.
- ^ Popper, Zoë A.; Michel, Gurvan; Hervé, Cécile; Domozych, David S.; Willats, William G.T.; Tuohy, Maria G.; Kloareg, Bernard; Stengel, Dagmar B. (2011). “Evolution and diversity of plant cell walls: From algae to flowering plants”. Annual Review of Plant Biology 62: 567–590. doi:10.1146/annurev-arplant-042110-103809. hdl:10379/6762. PMID 21351878.
- ^ Harper, Douglas. "eukaryotic". Online Etymology Dictionary.
- ^ Bonev, B; Cavalli, G (14 October 2016). “Organization and function of the 3D genome”. Nature Reviews Genetics 17 (11): 661–678. doi:10.1038/nrg.2016.112. hdl:2027.42/151884. PMID 27739532.
- ^ O'Connor, Clare (2008年). “Chromosome Segregation: The Role of Centromeres”. Nature Education. 2024年2月18日閲覧。 “eukar”
- ^ a b “Archean molecular fossils and the early rise of eukaryotes”. Science 285 (5430): 1033–1036. (August 1999). Bibcode: 1999Sci...285.1033B. doi:10.1126/science.285.5430.1033. PMID 10446042.
- ^ “The origin of the eukaryotic cell: a genomic investigation”. Proceedings of the National Academy of Sciences of the United States of America 99 (3): 1420–5. (February 2002). Bibcode: 2002PNAS...99.1420H. doi:10.1073/pnas.032658599. PMC 122206. PMID 11805300.
- ^ “Evolutionary Integration of Chloroplast Metabolism with the Metabolic Networks of the Cells”. Functional Genomics and Evolution of Photosynthetic Systems. Springer. (2011). p. 215. ISBN 978-94-007-1533-2. オリジナルの29 May 2016時点におけるアーカイブ。 2015年10月27日閲覧。
- ^ Endocytosis. Oxford University Press. (2001). p. vii. ISBN 978-0-19-963851-2
- ^ Stalder, Danièle; Gershlick, David C. (November 2020). “Direct trafficking pathways from the Golgi apparatus to the plasma membrane”. Seminars in Cell & Developmental Biology 107: 112–125. doi:10.1016/j.semcdb.2020.04.001. PMC 7152905. PMID 32317144.
- ^ “The nuclear envelope”. Cold Spring Harbor Perspectives in Biology 2 (3): a000539. (March 2010). doi:10.1101/cshperspect.a000539. PMC 2829960. PMID 20300205.
- ^ “Endoplasmic Reticulum (Rough and Smooth)”. British Society for Cell Biology. 2019年3月24日時点のオリジナルよりアーカイブ。2017年11月12日閲覧。
- ^ “Golgi Apparatus”. British Society for Cell Biology. 2017年11月13日時点のオリジナルよりアーカイブ。2017年11月12日閲覧。
- ^ “Lysosome”. British Society for Cell Biology. 2017年11月13日時点のオリジナルよりアーカイブ。2017年11月12日閲覧。
- ^ “Transcriptional profiling of lung cell populations in idiopathic pulmonary arterial hypertension”. Pulmonary Circulation 10 (1): 131–144. (July 1957). Bibcode: 1957SciAm.197a.131S. doi:10.1038/scientificamerican0757-131. PMC 7052475. PMID 32166015.
- ^ Fundamentals of Biochemistry (2nd ed.). John Wiley and Sons. (2006). pp. 547, 556. ISBN 978-0471214953
- ^ “Re: Are there eukaryotic cells without mitochondria?”. madsci.org (2006年5月1日). 2014年4月24日時点のオリジナルよりアーカイブ。2014年4月24日閲覧。
- ^ Zick, M; Rabl, R; Reichert, AS (January 2009). “Cristae formation-linking ultrastructure and function of mitochondria.”. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 1793 (1): 5–19. doi:10.1016/j.bbamcr.2008.06.013. PMID 18620004.
- ^ “28: The Origins of Life”. Molecular Biology of the Gene (Fourth ed.). Menlo Park, California: The Benjamin/Cummings Publishing Company, Inc.. (1988). p. 1154. ISBN 978-0-8053-9614-0
- ^ a b “A Eukaryote without a Mitochondrial Organelle”. Current Biology 26 (10): 1274–1284. (May 2016). doi:10.1016/j.cub.2016.03.053. PMID 27185558.
- ^ “Scientists Shocked To Discover Eukaryote With NO Mitochondria”. IFL Science (2016年5月13日). 2019年2月17日時点のオリジナルよりアーカイブ。2016年5月13日閲覧。
- ^ “Origin and Evolution of Plastids: Genomic View on the Unification and Diversity of Plastids”. The Structure and Function of Plastids. Advances in Photosynthesis and Respiration. 23. Springer Netherlands. (2006). pp. 75–102. doi:10.1007/978-1-4020-4061-0_4. ISBN 978-1-4020-4060-3
- ^ “Chloroplast DNA content in Dinophysis (Dinophyceae) from different cell cycle stages is consistent with kleptoplasty”. Environ. Microbiol. 10 (9): 2411–7. (September 2008). Bibcode: 2008EnvMi..10.2411M. doi:10.1111/j.1462-2920.2008.01666.x. PMID 18518896.
- ^ “Did some red alga-derived plastids evolve via kleptoplastidy? A hypothesis”. Biological Reviews of the Cambridge Philosophical Society 93 (1): 201–222. (February 2018). doi:10.1111/brv.12340. PMID 28544184.
- ^ Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter (2002-01-01). “Molecular Motors”. Molecular Biology of the Cell (4th ed.). New York: Garland Science. ISBN 978-0-8153-3218-3. オリジナルの8 March 2019時点におけるアーカイブ。 2023年4月6日閲覧。
- ^ “Motor Proteins”. Cold Spring Harbor Perspectives in Biology 10 (5): a021931. (May 2018). doi:10.1101/cshperspect.a021931. PMC 5932582. PMID 29716949.
- ^ “Prokaryotic motility structures”. Microbiology 149 (Pt 2): 295–304. (February 2003). doi:10.1099/mic.0.25948-0. PMID 12624192.
- ^ “Assembly and motility of eukaryotic cilia and flagella. Lessons from Chlamydomonas reinhardtii”. Plant Physiology 127 (4): 1500–7. (December 2001). doi:10.1104/pp.010807. PMC 1540183. PMID 11743094.
- ^ The centrosome and its role in the organization of microtubules. International Review of Cytology. 106. (1987). pp. 227–293. doi:10.1016/S0074-7696(08)61714-3. ISBN 978-0-12-364506-7. PMID 3294718
- ^ The Surprising Archaea: Discovering Another Domain of Life. Oxford: Oxford University Press. (2000). pp. 69–71. ISBN 978-0-19-511183-5
- ^ “The Structure and Functions of Xyloglucan”. Journal of Experimental Botany 40 (1): 1–11. (1989). doi:10.1093/jxb/40.1.1.
- ^ Hamilton, Matthew B. (2009). Population genetics. Wiley-Blackwell. p. 55. ISBN 978-1-4051-3277-0
- ^ Taylor, TN; Kerp, H; Hass, H (2005). “Life history biology of early land plants: Deciphering the gametophyte phase”. Proceedings of the National Academy of Sciences of the United States of America 102 (16): 5892–5897. doi:10.1073/pnas.0501985102. PMC 556298. PMID 15809414.
- ^ “Energetics and genetics across the prokaryote-eukaryote divide”. Biology Direct 6 (1): 35. (June 2011). doi:10.1186/1745-6150-6-35. PMC 3152533. PMID 21714941.
- ^ “The first sexual lineage and the relevance of facultative sex”. Journal of Molecular Evolution 48 (6): 779–783. (June 1999). Bibcode: 1999JMolE..48..779D. doi:10.1007/PL00013156. PMID 10229582.
- ^ a b “A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis”. Current Biology 15 (2): 185–191. (January 2005). doi:10.1016/j.cub.2005.01.003. PMID 15668177.
- ^ a b “An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis”. PLOS ONE 3 (8): e2879. (August 2007). Bibcode: 2008PLoSO...3.2879M. doi:10.1371/journal.pone.0002879. PMC 2488364. PMID 18663385.
- ^ “Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector”. Science 324 (5924): 265–268. (April 2009). Bibcode: 2009Sci...324..265A. doi:10.1126/science.1169464. PMC 2729066. PMID 19359589.
- ^ “The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms”. Proceedings: Biological Sciences 278 (1715): 2081–2090. (July 2011). doi:10.1098/rspb.2011.0289. PMC 3107637. PMID 21429931.
- ^ Patrick John Keeling; Yana Eglit (2023年11月21日), “Openly available illustrations as tools to describe eukaryotic microbial diversity” (英語), PLOS バイオロジー 21 (11): e3002395, doi:10.1371/JOURNAL.PBIO.3002395, ISSN 1544-9173, PMC 10662721, PMID 37988341, Wikidata Q123558544
- ^ “Taxonomic proposals for the classification of marine yeasts and other yeast-like fungi including the smuts”. Botanica Marina 23: 361–373. (1980).
- ^ Goldfuß (1818). “Ueber die Classification der Zoophyten [On the classification of zoophytes]” (ドイツ語). Isis, Oder, Encyclopädische Zeitung von Oken 2 (6): 1008–1019. オリジナルの24 March 2019時点におけるアーカイブ。 2019年3月15日閲覧。. From p. 1008: "Erste Klasse. Urthiere. Protozoa." (First class. Primordial animals. Protozoa.) [Note: each column of each page of this journal is numbered; there are two columns per page.]
- ^ “Not plants or animals: a brief history of the origin of Kingdoms Protozoa, Protista and Protoctista”. International Microbiology 2 (4): 207–221. (1999). PMID 10943416. オリジナルの14 June 2011時点におけるアーカイブ。.
- ^ a b “Protozoa, Protista, Protoctista: what's in a name?”. Journal of the History of Biology 22 (2): 277–305. (1989). doi:10.1007/BF00139515. PMID 11542176. オリジナルの4 February 2020時点におけるアーカイブ。 2020年2月4日閲覧。.
- ^ “New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms”. Science 163 (3863): 150–60. (January 1969). Bibcode: 1969Sci...163..150W. doi:10.1126/science.163.3863.150. PMID 5762760.
- ^ a b Sapp, J. (2005). “The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology”. Microbiol. Mol. Biol. Rev. 69 (2): 292-305. doi:10.1128/mmbr.69.2.292-305.2005.
- ^ a b c Katscher, F. (2004). “The History of the Terms Prokaryotes and Eukaryotes”. Protist 155: 257-263. doi:10.1078/143446104774199637.
- ^ Chatton, E. (1925). “Pansporella perplexa, amoebien à spores protégées parasite de daphnies”. Ann. Sci. Nat. Zool. 8: 5–84.
- ^ Lwoff, A. (1932). Recherches Biochimiques sur la Nutrition des Protozoaires. Le Pouvoir de Synthèse.. Masson
- ^ Copeland, H. (1938). “The kingdoms of organisms”. Quart. Rev. Biol. 13 (4): 383-420. doi:10.1086/394568.
- ^ Stanier & van Niel (1962). “The concept of a bacterium”. Arch. Mikrobiol. 42: 17–35. doi:10.1007/BF00425185.
- ^ Knoll, Andrew H. (1992). “The Early Evolution of Eukaryotes: A Geological Perspective”. Science 256 (5057): 622–627. Bibcode: 1992Sci...256..622K. doi:10.1126/science.1585174. PMID 1585174. "Eucarya, or eukaryotes"
- ^ Zaremba-Niedzwiedzka, Katarzyna; Caceres, Eva F.; Saw, Jimmy H.; Bäckström, Disa; Juzokaite, Lina; Vancaester, Emmelien; Seitz, Kiley W.; Anantharaman, Karthik et al. (2017-01). “Asgard archaea illuminate the origin of eukaryotic cellular complexity” (英語). Nature 541 (7637): 353–358. doi:10.1038/nature21031. ISSN 0028-0836.
- ^ “Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista”. Proceedings: Biological Sciences 283 (1823): 20152802. (January 2016). doi:10.1098/rspb.2015.2802. PMC 4795036. PMID 26817772.
- ^ a b Brown, Matthew W.; Heiss, Aaron A.; Kamikawa, Ryoma; Inagaki, Yuji; Yabuki, Akinori; Tice, Alexander K; Shiratori, Takashi; Ishida, Ken-Ichiro et al. (2018-01-19). “Phylogenomics Places Orphan Protistan Lineages in a Novel Eukaryotic Super-Group”. Genome Biology and Evolution 10 (2): 427–433. doi:10.1093/gbe/evy014. PMC 5793813. PMID 29360967.
- ^ “Picozoa are archaeplastids without plastid”. Nature Communications 12 (1): 6651. (2021). doi:10.1038/s41467-021-26918-0. PMC 8599508. PMID 34789758. オリジナルの2 February 2024時点におけるアーカイブ。 2021年12月20日閲覧。.
- ^ “Picozoa are archaeplastids without plastid”. Nature Communications 12 (1): 6651. (2021). doi:10.1038/s41467-021-26918-0. PMC 8599508. PMID 34789758.
- ^ “Microbial predators form a new supergroup of eukaryotes”. Nature 612 (7941): 714–719. (December 2022). doi:10.1038/s41586-022-05511-5. PMID 36477531.
- ^ a b “The role of symbiosis in eukaryotic evolution”. Origins and Evolution of Life: An astrobiological perspective. Cambridge: Cambridge University Press. (2011). pp. 326–339. ISBN 978-0-521-76131-4. オリジナルの24 March 2019時点におけるアーカイブ。 2017年8月27日閲覧。
- ^ “Origin and Early Evolution of the Eukaryotic Cell”. Annual Review of Microbiology 75 (1): 631–647. (October 2021). doi:10.1146/annurev-micro-090817-062213. PMID 34343017.
- ^ “Concepts of the last eukaryotic common ancestor”. Nature Ecology & Evolution 3 (3): 338–344. (March 2019). Bibcode: 2019NatEE...3..338O. doi:10.1038/s41559-019-0796-3. hdl:10261/201794. PMID 30778187.
- ^ “Predatory protists”. Current Biology 30 (10): R510–R516. (May 2020). doi:10.1016/j.cub.2020.03.052. PMID 32428491.
- ^ “A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids”. Nature Communications 12 (1): 1879. (March 2021). Bibcode: 2021NatCo..12.1879S. doi:10.1038/s41467-021-22044-z. PMC 7994803. PMID 33767194.
- ^ Koumandou, V. Lila; Wickstead, Bill; Ginger, Michael L.; van der Giezen, Mark; Dacks, Joel B.; Field, Mark C. (2013). “Molecular paleontology and complexity in the last eukaryotic common ancestor”. Critical Reviews in Biochemistry and Molecular Biology 48 (4): 373–396. doi:10.3109/10409238.2013.821444. PMC 3791482. PMID 23895660.
- ^ “Actin cytoskeleton and complex cell architecture in an Asgard archaean”. Nature 613 (7943): 332–339. (2023). Bibcode: 2023Natur.613..332R. doi:10.1038/s41586-022-05550-y. hdl:20.500.11850/589210. PMC 9834061. PMID 36544020.
- ^ Martin, William; Müller, Miklós (1998-03). “The hydrogen hypothesis for the first eukaryote” (英語). Nature 392 (6671): 37–41. doi:10.1038/32096. ISSN 0028-0836.
- ^ Spang, Anja; Stairs, Courtney W.; Dombrowski, Nina; Eme, Laura; Lombard, Jonathan; Caceres, Eva F.; Greening, Chris; Baker, Brett J. et al. (2019-07). “Proposal of the reverse flow model for the origin of the eukaryotic cell based on comparative analyses of Asgard archaeal metabolism” (英語). Nature Microbiology 4 (7): 1138–1148. doi:10.1038/s41564-019-0406-9. ISSN 2058-5276.
- ^ Imachi, Hiroyuki; Nobu, Masaru K.; Nakahara, Nozomi; Morono, Yuki; Ogawara, Miyuki; Takaki, Yoshihiro; Takano, Yoshinori; Uematsu, Katsuyuki et al. (2020-01-23). “Isolation of an archaeon at the prokaryote–eukaryote interface” (英語). Nature 577 (7791): 519–525. doi:10.1038/s41586-019-1916-6. ISSN 0028-0836. PMC 7015854. PMID 31942073.
- ^ López-García, Purificación; Moreira, David (2020-05). “The Syntrophy hypothesis for the origin of eukaryotes revisited” (英語). Nature Microbiology 5 (5): 655–667. doi:10.1038/s41564-020-0710-4. ISSN 2058-5276.
- ^ Moreira, David; López-García, Purificación (1998-11). “Symbiosis Between Methanogenic Archaea and δ-Proteobacteria as the Origin of Eukaryotes: The Syntrophic Hypothesis” (英語). Journal of Molecular Evolution 47 (5): 517–530. doi:10.1007/PL00006408. ISSN 0022-2844.
- ^ Fuerst, John A.; Sagulenko, Evgeny (2011-06). “Beyond the bacterium: planctomycetes challenge our concepts of microbial structure and function” (英語). Nature Reviews Microbiology 9 (6): 403–413. doi:10.1038/nrmicro2578. ISSN 1740-1526.
- ^ Roger, Andrew J.; Muñoz-Gómez, Sergio A.; Kamikawa, Ryoma (2017-11). “The Origin and Diversification of Mitochondria”. Current Biology 27 (21): R1177–R1192. doi:10.1016/j.cub.2017.09.015. ISSN 0960-9822.
- ^ a b Hoshino, Yosuke; Gaucher, Eric A. (2021-06-22). “Evolution of bacterial steroid biosynthesis and its impact on eukaryogenesis” (英語). Proceedings of the National Academy of Sciences 118 (25). doi:10.1073/pnas.2101276118. ISSN 0027-8424. PMC 8237579. PMID 34131078.
- ^ a b Knoll, A.h; Javaux, E.j; Hewitt, D; Cohen, P (2006-06-29). “Eukaryotic organisms in Proterozoic oceans”. Philosophical Transactions of the Royal Society B: Biological Sciences 361 (1470): 1023–1038. doi:10.1098/rstb.2006.1843. PMC 1578724. PMID 16754612.
- ^ Müller, Miklós; Mentel, Marek; van Hellemond, Jaap J.; Henze, Katrin; Woehle, Christian; Gould, Sven B.; Yu, Re-Young; van der Giezen, Mark et al. (2012-06). “Biochemistry and Evolution of Anaerobic Energy Metabolism in Eukaryotes” (英語). Microbiology and Molecular Biology Reviews 76 (2): 444–495. doi:10.1128/MMBR.05024-11. ISSN 1092-2172. PMC 3372258. PMID 22688819.
- ^ Martin, William F.; Tielens, Aloysius G. M.; Mentel, Marek (2020-12-07). Mitochondria and Anaerobic Energy Metabolism in Eukaryotes: Biochemistry and Evolution. De Gruyter. doi:10.1515/9783110612417. ISBN 978-3-11-061241-7
- ^ Catling, David C.; Zahnle, Kevin J. (2020-02). “The Archean atmosphere” (英語). Science Advances 6 (9): eaax1420. doi:10.1126/sciadv.aax1420. ISSN 2375-2548. PMC 7043912. PMID 32133393.
- ^ “Mass extinctions: the microbes strike back”. New Scientist: 40–43. (9 February 2008). オリジナルの8 July 2008時点におけるアーカイブ。 2017年8月27日閲覧。.
- ^ Brocks, J. J. (1999-08-13). “Archean Molecular Fossils and the Early Rise of Eukaryotes”. Science 285 (5430): 1033–1036. doi:10.1126/science.285.5430.1033.
- ^ Waldbauer, Jacob R.; Sherman, Laura S.; Sumner, Dawn Y.; Summons, Roger E. (2009-03). “Late Archean molecular fossils from the Transvaal Supergroup record the antiquity of microbial diversity and aerobiosis” (英語). Precambrian Research 169 (1-4): 28–47. doi:10.1016/j.precamres.2008.10.011.
- ^ Brocks, Jochen J; Buick, Roger; Summons, Roger E; Logan, Graham A (2003-11). “A reconstruction of Archean biological diversity based on molecular fossils from the 2.78 to 2.45 billion-year-old Mount Bruce Supergroup, Hamersley Basin, Western Australia” (英語). Geochimica et Cosmochimica Acta 67 (22): 4321–4335. doi:10.1016/S0016-7037(03)00209-6.
- ^ French, Katherine L.; Hallmann, Christian; Hope, Janet M.; Schoon, Petra L.; Zumberge, J. Alex; Hoshino, Yosuke; Peters, Carl A.; George, Simon C. et al. (2015-05-12). “Reappraisal of hydrocarbon biomarkers in Archean rocks”. Proceedings of the National Academy of Sciences 112 (19): 5915–5920. doi:10.1073/pnas.1419563112. PMC 4434754. PMID 25918387.
- ^ “Reappraisal of hydrocarbon biomarkers in Archean rocks”. Proceedings of the National Academy of Sciences of the United States of America 112 (19): 5915–5920. (May 2015). Bibcode: 2015PNAS..112.5915F. doi:10.1073/pnas.1419563112. PMC 4434754. PMID 25918387.
- ^ Rasmussen, Birger; Fletcher, Ian R.; Brocks, Jochen J.; Kilburn, Matt R. (2008-10). “Reassessing the first appearance of eukaryotes and cyanobacteria” (英語). Nature 455 (7216): 1101–1104. doi:10.1038/nature07381. ISSN 0028-0836.
- ^ “The rise of algae in Cryogenian oceans and the emergence of animals”. Nature 548 (7669): 578–581. (August 2017). Bibcode: 2017Natur.548..578B. doi:10.1038/nature23457. PMID 28813409.
- ^ Hoshino, Yosuke; Poshibaeva, Aleksandra; Meredith, William; Snape, Colin; Poshibaev, Vladimir; Versteegh, Gerard J. M.; Kuznetsov, Nikolay; Leider, Arne et al. (2017-09). “Cryogenian evolution of stigmasteroid biosynthesis” (英語). Science Advances 3 (9): e1700887. doi:10.1126/sciadv.1700887. ISSN 2375-2548. PMC 5606710. PMID 28948220.
- ^ Brocks, Jochen J.; Jarrett, Amber J. M.; Sirantoine, Eva; Hallmann, Christian; Hoshino, Yosuke; Liyanage, Tharika (2017-08). “The rise of algae in Cryogenian oceans and the emergence of animals” (英語). Nature 548 (7669): 578–581. doi:10.1038/nature23457. ISSN 1476-4687.
- ^ “Paleoproterozoic sterol biosynthesis and the rise of oxygen”. Nature 543 (7645): 420–423. (March 2017). Bibcode: 2017Natur.543..420G. doi:10.1038/nature21412. hdl:1721.1/128450. PMID 28264195.
- ^ “Sterol Synthesis in Diverse Bacteria”. Frontiers in Microbiology 7: 990. (2016-06-24). doi:10.3389/fmicb.2016.00990. PMC 4919349. PMID 27446030.
- ^ “Evolution of bacterial steroid biosynthesis and its impact on eukaryogenesis”. Proceedings of the National Academy of Sciences of the United States of America 118 (25): e2101276118. (June 2021). Bibcode: 2021PNAS..11801276H. doi:10.1073/pnas.2101276118. PMC 8237579. PMID 34131078.
- ^ Brocks, Jochen J.; Nettersheim, Benjamin J.; Adam, Pierre; Schaeffer, Philippe; Jarrett, Amber J. M.; Güneli, Nur; Liyanage, Tharika; van Maldegem, Lennart M. et al. (2023-06-22). “Lost world of complex life and the late rise of the eukaryotic crown” (英語). Nature 618 (7966): 767–773. doi:10.1038/s41586-023-06170-w. ISSN 0028-0836.
- ^ Albani, Abderrazak El; Bengtson, Stefan; Canfield, Donald E.; Bekker, Andrey; Macchiarelli, Roberto; Mazurier, Arnaud; Hammarlund, Emma U.; Boulvais, Philippe et al. (2010-07). “Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago” (英語). Nature 466 (7302): 100–104. doi:10.1038/nature09166. ISSN 1476-4687.
- ^ Strassert, Jürgen F. H.; Irisarri, Iker; Williams, Tom A.; Burki, Fabien (2021-03-25). “A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids” (英語). Nature Communications 12 (1): 1879. doi:10.1038/s41467-021-22044-z. ISSN 2041-1723. PMC 7994803. PMID 33767194.
- ^ Parfrey, L. W.; Lahr, D. J. G.; Knoll, A. H.; Katz, L. A. (2011-08-16). “Estimating the timing of early eukaryotic diversification with multigene molecular clocks” (英語). Proceedings of the National Academy of Sciences 108 (33): 13624–13629. doi:10.1073/pnas.1110633108. ISSN 0027-8424. PMC 3158185. PMID 21810989.
- ^ “Tracking the rise of eukaryotes to ecological dominance with zinc isotopes”. Geobiology 16 (4): 341–352. (June 2018). Bibcode: 2018Gbio...16..341I. doi:10.1111/gbi.12289. PMID 29869832.
- ^ Miao, L.; Yin, Z.; Knoll, A. H.; Qu, Y.; Zhu, M. (2024). “1.63-billion-year-old multicellular eukaryotes from the Chuanlinggou Formation in North China”. Science Advances 10 (4): eadk3208. doi:10.1126/sciadv.adk3208. PMC 10807817. PMID 38266082.
- ^ “Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan”. Science 257 (5067): 232–5. (July 1992). Bibcode: 1992Sci...257..232H. doi:10.1126/science.1631544. PMID 1631544.
- ^ “Eukaryotic organisms in Proterozoic oceans”. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 361 (1470): 1023–1038. (June 2006). doi:10.1098/rstb.2006.1843. PMC 1578724. PMID 16754612.
- ^ a b c “Problematic urn-shaped fossils from a Paleoproterozoic (2.2 Ga) paleosol in South Africa.”. Precambrian Research 235: 71–87. (2013). Bibcode: 2013PreR..235...71R. doi:10.1016/j.precamres.2013.05.015.
- ^ “Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago”. Nature 466 (7302): 100–104. (July 2010). Bibcode: 2010Natur.466..100A. doi:10.1038/nature09166. PMID 20596019.
- ^ El Albani, Abderrazak (2023). “A search for life in Palaeoproterozoic marine sediments using Zn isotopes and geochemistry”. Earth and Planetary Science Letters 623: 118169. Bibcode: 2023E&PSL.61218169E. doi:10.1016/j.epsl.2023.118169.
- ^ Ossa Ossa, Frantz; Pons, Marie-Laure; Bekker, Andrey; Hofmann, Axel; Poulton, Simon W. et al. (2023). “Zinc enrichment and isotopic fractionation in a marine habitat of the c. 2.1 Ga Francevillian Group: A signature of zinc utilization by eukaryotes?”. Earth and Planetary Science Letters 611: 118147. Bibcode: 2023E&PSL.61118147O. doi:10.1016/j.epsl.2023.118147.
- ^ Fakhraee, Mojtaba; Tarhan, Lidya G.; Reinhard, Christopher T.; Crowe, Sean A.; Lyons, Timothy W.; Planavsky, Noah J. (May 2023). “Earth's surface oxygenation and the rise of eukaryotic life: Relationships to the Lomagundi positive carbon isotope excursion revisited” (英語). Earth-Science Reviews 240: 104398. Bibcode: 2023ESRv..24004398F. doi:10.1016/j.earscirev.2023.104398. オリジナルの2 February 2024時点におけるアーカイブ。 2023年6月6日閲覧。.
- ^ “The controversial "Cambrian" fossils of the Vindhyan are real but more than a billion years older”. Proceedings of the National Academy of Sciences of the United States of America 106 (19): 7729–7734. (May 2009). Bibcode: 2009PNAS..106.7729B. doi:10.1073/pnas.0812460106. PMC 2683128. PMID 19416859.
参考文献
- [Adl(2005)] Adl, Sina M.; Simpson, Alastair G. B.; Farmer, Mark A.; Andersen, Robert A.; Anderson, O. Roger; Barta, John R.; Bowser, Samuel S.; Brugerolle, Guy et al. (2005-10). “The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists” (英語). The Journal of Eukaryotic Microbiology 52 (5): 399–451. doi:10.1111/j.1550-7408.2005.00053.x. ISSN 1066-5234.
- [Adl(2012)] Adl, Sina M.; Simpson, Alastair G. B.; Lane, Christopher E.; Lukeš, Julius; Bass, David; Bowser, Samuel S.; Brown, Matthew W.; Burki, Fabien et al. (2012-09). “The Revised Classification of Eukaryotes” (英語). Journal of Eukaryotic Microbiology 59 (5): 429–514. doi:10.1111/j.1550-7408.2012.00644.x. PMC 3483872. PMID 23020233.
- [Adl(2019)] Adl, Sina M.; Bass, David; Lane, Christopher E.; Lukeš, Julius; Schoch, Conrad L.; Smirnov, Alexey; Agatha, Sabine; Berney, Cedric et al. (2019). “Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes” (英語). Journal of Eukaryotic Microbiology 66. doi:10.1111/jeu.12691. ISSN 1066-5234. PMC 6492006. PMID 30257078.
- [矢﨑(2020)] 矢﨑裕規; 島野智之『真核生物の高次分類体系の改訂―Adl et al. (2019)について―』日本動物分類学会、2020年2月29日。doi:10.19004/taxa.48.0_71。2020年3月24日閲覧。