
「卵菌」の版間の差分
m Category:不等毛類を除去; Category:ストラメノパイルを追加 (HotCat使用) |
「日和見的な寄生菌」に日和見感染へのリンクを追加 |
||
| (2人の利用者による、間の4版が非表示) | |||
| 1行目: | 1行目: | ||
{{生物分類表 |
{{生物分類表 |
||
| 名称 = 卵菌 |
| 名称 = 卵菌 |
||
| 色 = |
| 色 = 原生生物界 |
||
| 画像 = [[画像: |
| 画像 = [[画像:Achlya oospore.jpg|250px]] |
||
| 画像キャプション = |
| 画像キャプション = '''1'''. ワタカビ属(ミズカビ目)の菌糸と卵胞子 |
||
| ドメイン = [[真核生物]] |
| ドメイン = [[真核生物]] {{Sname||Eukaryota}} |
||
| 界 = [[ |
| 上界階級なし = [[ディアフォレティケス]] {{Sname||Diaphoretickes}} |
||
| 界階級なし = [[SARスーパーグループ]] {{Sname||SAR supergroup}} |
|||
| 門 = '''卵菌門''' Oomycota |
|||
| 亜界階級なし = [[ストラメノパイル]] {{Sname||Stramenopiles}} |
|||
| 上門階級なし = {{Sname||Gyrista}} |
|||
| 門 = [[偽菌類|偽菌門]] {{sname||Pseudofungi}}<br /><small>または</small>'''卵菌門''' {{Sname||Oomycota}} |
|||
| 綱 = '''卵菌綱''' {{sname||Oomycetes}} |
|||
| 学名 = |
|||
{{Sname||Oomycota}} {{AUY|Arx|1967|bio=bot}}<br /> |
|||
{{Sname||Oomycetes}} {{AUY|G. Winter|1880|bio=bot}} |
|||
| シノニム = |
|||
* {{Sname||Peronosporomycetes}} {{AUY|Locq.|1974|bio=bot}}(広義) |
|||
| 英名 = oomycete |
|||
| 下位分類名 = 下位分類 |
| 下位分類名 = 下位分類 |
||
| 下位分類 = {{ |
| 下位分類 = {{center|[[#system|本文参照]]}} |
||
}} |
}} |
||
'''卵菌'''(らんきん、{{lang-en-short|oomycetes}})とは、[[ストラメノパイル]]に属する吸収栄養性([[細胞膜]]を通して低分子[[有機物]]を吸収する)の[[原生生物]]の一群、またはこれに属する生物のことであり、その形態や生活様式は[[菌類]]に似ているが、系統的には全く異なる。微小な単細胞性のものから、発達した[[菌糸]]を形成するものまであり、菌糸は基本的に隔壁を欠く(図1)。[[セルロース]]を含む[[細胞壁]]をもつ。多くは、2本の不等[[鞭毛]]をもつ[[遊走子]]による[[無性生殖]]を行い、配偶子嚢接合による[[有性生殖]]を行う(図1)。栄養体は[[核相|複相]]([[染色体]]を2セットもつ)であり、配偶子嚢内で[[減数分裂]]を行う。海や淡水、陸上に広く分布しており、腐生性(生物遺体など生きていない有機物を分解して栄養を吸収する)、または[[陸上植物]]や[[藻類]]、[[動物]]などに寄生する。植物病原菌となるものが多く知られており、[[ジャガイモエキビョウキン]]([[ツユカビ目]])は19世紀に[[アイルランド]]で[[ジャガイモ飢饉]]を引き起こし、多数の餓死者と[[北米]]への移民の原因となった。 |
|||
100属1,700種ほどが知られる大きなグループであり、分類学的には、'''卵菌綱'''([[学名]]: {{Sname||Oomycetes}})にまとめられることが多いが、独自の'''卵菌門'''(学名: {{Sname||Oomycota}})にまとめて複数の綱に分けることもある。[[菌類]]に似ているため、古くは[[菌類]]の中の[[藻菌類]]や[[鞭毛菌類]]に分類されていた。しかし卵菌と菌類は系統的に非常に離れており、この類似性は[[収斂進化]]によるものである。 |
|||
'''卵菌'''(らんきん)は、[[不等毛類]]に属する[[原生生物]]の一つ。近縁の[[サカゲカビ|サカゲツボカビ類]]とともに[[菌類]]様の外見を持つものが多い。 |
|||
== 特徴 == |
|||
化石記録としては、[[石炭紀]]の地層から[[ヒカゲノカズラ類]]の内部寄生者、{{Snamei||Combresomyces}} 属の化石が発見されているほか<ref>{{cite journal|author=Nora DOTZLER, Michael KRINGS, Reinhard AGERER, |
|||
=== 体制 === |
|||
Jean GALTIER, Thomas N. TAYLOR|title=Combresomyces cornifer gen. sp. nov., an endophytic peronosporomycete in Lepidodendron from the Carboniferous of central France|}}</ref>、[[白亜紀]]の[[琥珀]]からも卵菌類と考えられる構造が見つかっている<ref>{{cite web|url=http://www.ucmp.berkeley.edu/chromista/oomycota.html|title=Introduction to the Oomycota|accessdate=2012-09-}}</ref>。 |
|||
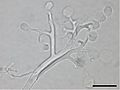
菌体の体制はさまざまであり、単細胞で[[全実性]](全体が[[胞子嚢|遊走子嚢]]など生殖器官になる)のものから、発達した[[菌糸]]を形成するもの(下図2)まである<ref name="Webster2007">{{Cite book|author=Webster, J. & Weber, R. W. S.|year=2007|chapter=Straminipila: Oomycota|editor=|title=Introduction to Fungi 3rd Edition|publisher=Cambridge University Press|isbn=978-0521014830|pages=75–126}}</ref><ref name="Beakes2014">{{cite book|author=Beakes, G. W., Honda, D. & Thines, M.|year=2014|chapter=Systematics of the Straminipila: Labyrinthulomycota, Hyphochytriomycota, and Oomycota|editor=McLaughlin, D. J. & Spatafora, J. W.|title=THE MYCOTA, volume 7A. Systematics and Evolution Part A|publisher=Springer|doi=10.1007/978-3-642-55318-9_3|pages=39-97}}</ref><ref name="Beakes2017">{{cite book|author=Beakes, G. W. & Thines, M.|year=2017|chapter=Hyphochytriomycota and Oomycota|editor=Archibald, J. M., Simpson, A. G. B. & Slamovits, C. H.|title=Handbook of the Protists|publisher=Springer|isbn=978-3319281476|pages=435-505}}</ref><ref name="徳増2005" />。全実性のものでは、単純な球状の種から、菌糸状に細長いものや、分岐・分節化した種まである<ref name="Webster2007" /><ref name="Beakes2017" />。卵菌の中で初期分岐群は全実性・寄生性であるが、ミズカビ目やツユカビ目など派生的な群では発達した菌糸をもつものが多い<ref name="Beakes2017" /><ref name="Beakes2009">{{cite web|title=The evolutionary phylogeny of Oomycetes—insights gained from studies of holocarpic parasites of algae and invertebrates|url=http://media.wiley.com/product_data/excerpt/76/04702556/0470255676.pdf|accessdate=2012-10-25}}</ref><ref name="Beakes2012">{{cite journal|author=Gordon W. Beakes & Sally L. Glockling & Satoshi Sekimoto|title=The evolutionary phylogeny of the oomycete “fungi”|journal=Protoplasma|year=2012|volume=249|pages=3–19|doi=10.1007/s00709-011-0269-2}}</ref>。ただし派生的な群の中にも、全実性・寄生性である種が含まれる。菌糸の太さはさまざまであり、直径2[[マイクロメートル]] (µm) 程度のものから、150 µm に達するものまである<ref name="Beakes2017" />。菌糸は先端成長を行い、ときに分枝し、ふつう隔壁を欠く'''無隔菌糸'''であるが(下図2)、古くなった部分や生殖器を区切る隔壁が形成されることもある<ref name="Webster2007" /><ref name="Beakes2017" />。菌糸性で絶対寄生性の種の多くは、宿主細胞間に菌糸を伸ばし、宿主細胞内に[[吸器]]を挿入する<ref name="Webster2007" /><ref name="Beakes2017" />(下図2c)。 |
|||
{{multiple image |
|||
== 系統 == |
|||
| total_width = 600 |
|||
かつてはツボカビ類・接合菌類などとともに[[鞭毛菌門|鞭毛菌]]類あるいは[[藻菌類]]と呼ばれた。これは、菌類が退化した藻類であるとする、菌類の藻類起源説に立ち、藻類に近いものとしてこれらの類が考えられたことに由来する。特に、卵菌類と[[黄緑藻類]]の[[フシナシミドロ]]の類似が指摘されたこともある。その後、細胞壁や鞭毛の構造が明らかに異なることから、菌類が独立の界と見なされるようになり、藻類起源説は力を失った。しかし現在でも[[植物病理学]]など応用分野で便宜的に藻菌や鞭毛菌と呼ばれることがある。 |
|||
| align = center |
|||
| caption_align = left |
|||
| image1 = Kreeftenpest.jpg |
|||
| caption1 = '''2a'''. {{Snamei||Aphanomyces}}(ミズカビ目)の菌糸と胞子 |
|||
| image2 = Pythium_myriotylum.jpg |
|||
| caption2 = '''2b'''. {{Snamei||Pythium}}(ツユカビ目)の菌糸と胞子嚢 |
|||
| image3 = Bremia lactucae haustoria.jpg |
|||
| caption3 = '''2c'''. 宿主植物細胞間隙の {{Snamei||Bremia}}(ツユカビ目)の菌糸: 隣接する植物細胞内に[[吸器]]を挿入している |
|||
}} |
|||
菌体はふつう[[細胞壁]]に包まれるが、寄生性の初期分岐群の中には、栄養体(通常時の体)が宿主内で細胞壁を欠く多核細胞([[変形体]])として生きている例もある<ref name="Beakes2014" /><ref name="Beakes2017" />。卵菌の細胞壁はおもに[[セルロース]]と[[β-グルカン]]からなる。ただし一部の種からは[[キチン]]も報告されており、また[[ゲノム]]の調査からは全ての卵菌類がキチン合成酵素をもつことが示唆されている<ref name="Klinter2019">{{Cite journal|author=Klinter, S., Bulone, V. & Arvestad, L.|year=2019|title=Diversity and evolution of chitin synthases in oomycetes (Straminipila: Oomycota)|journal=Molecular Phylogenetics and Evolution|volume=139|issue=|pages=106558|doi=10.1016/j.ympev.2019.106558}}</ref>。フシミズカビ目では、菌糸のくびれ部分にキチンを含むセルリン顆粒が存在することがある<ref name="Beakes2017" />。 |
|||
先にも述べたように、この類はかつては菌類と見なされていたが、現在は別系統と考えられている。鞭毛の構造等から、褐藻や珪藻などの不等毛藻類などと共通の系統に属するものと見なされ、[[ストラメノパイル]]などの群に属させる。この群にはいくつかの藻類の他に、菌類と見なされたこともあるサカゲカビ類や[[ラビリンチュラ]]類も含まれている。しかし、それらと卵菌類の系統関係については明らかではない。特にサカゲカビ類はよく似た部分が多いが、近縁であるとの判断は必ずしも出されていない。いずれにせよ、藻類に近縁であるからといって、これらの菌類的生物が藻類に由来したものとは考えられていない。 |
|||
=== 細胞 === |
|||
卵菌と[[サカゲカビ]]はいずれも菌類的生物でありながら、菌類とは系統を異にする。この二者は、系統的には近いものとの判断から、これを一つにまとめる考えもある。この場合、二つをまとめる名称として、二毛菌という名称を使う。 |
|||
卵菌の栄養体は、基本的に多数の[[細胞核|核]]を含む'''多核性''' (coenocytic) である<ref name="Webster2007" /><ref name="Beakes2014" /><ref name="Beakes2017" />。[[核分裂]]は核膜が維持される閉鎖型であり、核内に[[紡錘体]]が形成され、両極には[[中心体]]が存在する<ref name="Webster2007" /><ref name="Beakes2017" />。 |
|||
栄養体中央はふつう[[液胞]]で占められており、[[細胞核|核]]や他の細胞小器官を含む細胞質は周縁部に位置する<ref name="Beakes2017" />。[[ミトコンドリア]]のクリステは管状である<ref name="Webster2007" /><ref name="Beakes2017" />。多くの[[菌類]]とは異なり、典型的な[[ゴルジ体]]をもつ<ref name="Webster2007" /><ref name="Beakes2017" />。貯蔵多糖は[[リン酸化]]した水溶性のβ-1,3[[グルカン]]であり、'''マイコラミナリン''' (mycolaminarin) とよばれる<ref name="Webster2007" />。マイコラミナリンは小胞に包まれてときに層状構造を示し、この小胞は dense-body vesicle または fingerprint vacuole ともよばれる<ref name="Webster2007" />。菌類に一般的な貯蔵多糖である[[グリコーゲン]]顆粒は、卵菌には見られない<ref name="Webster2007" />。また、[[ポリリン酸]]体を欠く<ref name="Webster2007" />。[[アミノ酸]]の[[リシン]]合成経路は[[ジアミノピメリン酸]]経路(DAP経路)であり、菌類に見られる[[α-アミノアジピン酸]]経路(AAA経路)とは異なる<ref name="Webster2007" />。 |
|||
[[ファイル:Water mold Mizukabi colony.jpg|260px|thumb|麻の実から育ったミズカビ類のコロニー<br>ほとんどは''Achlya'' sp.]] |
|||
== |
=== 無性生殖 === |
||
卵菌は、基本的に'''[[遊走子]]'''([[鞭毛]]をもつ胞子)'''による[[無性生殖]]を行う'''(下図3a)。菌体の全体または一部が遊走子嚢となり、[[細胞壁]]の一部が溶解することで遊走子を放出する<ref name="Beakes2017" /><ref name="徳増2005" />。ただし鞭毛を欠く不動胞子 (aplanospore) を放出することもあり、また鞭毛形成能を完全に欠いている種もいる<ref name="Beakes2017" />。遊走子は好適な環境に着生すると、再び菌体へと成長する<ref name="Webster2007" /><ref name="Beakes2017" />。[[全実性]]で寄生性の種では、放出される遊走子の通り道となる逸出管を形成することが多い<ref name="Beakes2017" />。[[菌糸]]を形成するものでは、ふつう菌糸先端が隔壁によって区切られて遊走子嚢を形成する<ref name="Beakes2017" />(下図3b左)。[[シロサビキン属]]([[シロサビキン目]])では、遊走子嚢が鎖状に連なって形成される<ref name="Beakes2017" />(下図3b中)。[[ツユカビ属]]([[ツユカビ目]])などでは、有限成長する分枝した遊走子嚢柄 (sporangiophore) の枝先に遊走子嚢をつける<ref name="Beakes2017" />(下図3b右, 3c)。 |
|||
栄養体は隔壁のない単[[細胞核|核]]あるいは[[多核体]]の[[葉状体]]で、多くの場合[[菌類]]の[[菌糸|菌糸体]]によく似た形態に[[収斂進化]]を起こしている。特に隔壁を欠いている点は菌類の中でも[[ツボカビ]]類や[[接合菌]]と類似している。他の卵菌に寄生するフクロカビモドキなどの葉状体は単純な袋状で菌糸状の形はとらない。葉状体は動植物遺体上で有機物を分解したり、他の[[生物]]に[[寄生]]して生活する。[[細胞壁]]の主成分は[[セルロース]]で、菌類([[キチン]])と異なる。 |
|||
卵菌の初期分岐群やミズカビ亜綱では、[[遊走子]]形成は遠心的であり、また細胞分裂が完了したのちに[[鞭毛]]形成が起こる<ref name="Beakes2017" />。それに対してツユカビ亜綱では、遊走子形成は求心的であり、また遊走子の分化と鞭毛形成は同時に起こる<ref name="Beakes2017" />。一部の卵菌では、遊走子嚢から放出された遊走子または原形質が包嚢 (vesicle) で包まれ、後者の場合はこの中で遊走子への分化が起こる<ref name="Beakes2017" />。陸上植物に寄生する種では、しばしば遊走子嚢が切り離されて風や雨で散布され、水の存在によって散布先で遊走子を放出するが、遊走子段階を省略して直接発芽することも多い<ref name="Webster2007" /><ref name="Beakes2017" />(下図3b右, 6)。このように直接発芽する遊走子嚢は、[[分生子]] (conidium) ともよばれる<ref name="Beakes2014" /><ref name="Beakes2017" />。 |
|||
遊走細胞は同じ[[不等毛類]]の[[褐藻]]などと同様で、[[マスチゴネマ]]を持つ羽型[[鞭毛]]と鞭型鞭毛をセットで持つ。葉状体は[[複相]](2n)で、[[無性生殖]]に際しては菌糸様の葉状体の先端に[[胞子嚢]]を形成し、この内部の原形質が分割されて多数の[[遊走子]]を生じ、これが泳ぎ出て[[宿主]]に到達する。葉状体に形成された一次型遊走子はクロミスタ界の中でも[[真眼点藻類]]の遊走細胞と同様に2本の鞭毛を細胞の前端に持つが、一次型遊走子がシスト化してから脱皮して生じる二次型遊走子は褐藻などと同様にソラマメ型の細胞の側面から前方に羽型鞭毛が、後方に鞭型鞭毛が伸びる。 |
|||
{{multiple image |
|||
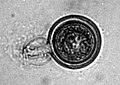
[[ファイル:Achlya oospore.jpg|right|280px|thumb|ワタカビ属の1種(''Achlya'' sp.)の卵胞子]] |
|||
| total_width = 800 |
|||
[[有性生殖]]は、[[配偶子]]が独立せず、配偶子嚢の状態で接合する'''[[配偶子嚢接合]]'''という形を取る。まず葉状体に多核の[[造精器]]と[[生卵器]]が互いに接して形成され、それぞれの内部で[[減数分裂]]が行われる。造精器から生卵器に受精管が伸び、単相(n)の[[配偶子]]核が移送されると生卵器内に受精によって生じた厚壁の卵胞子が1-40個形成される。この卵胞子を形成する有性生殖により卵菌と呼ばれる。卵胞子が休眠後発芽すると遊走子嚢が突出し、そこから多数の遊走子が放出される。 |
|||
| align = center |
|||
| caption_align = left |
|||
| image1 = Phytophthora_capsici_zoospores.webm |
|||
| caption1 = '''3a'''. [[エキビョウキン|エキビョウキン属]]の遊走子 |
|||
| image2 = 07 07 asexual structures, Saprolegniales, Albuginales, Peronosporales, Oomycota (M. Piepenbring).png |
|||
| caption2 = '''3b'''. ミズカビ目 (左)、シロサビキン目 (中)、ツユカビ目 (右) の遊走子嚢: sporangium = 遊走子嚢、sporangiophore = 遊走子嚢柄、zoospore = 遊走子、spore = 胞子、sporophore = 担胞子体、conidium = 分生子、stoma = 気孔 |
|||
| image3 = Zoospore_release.jpg |
|||
| caption3 = '''3c'''. エキビョウキン属(ツユカビ目): 遊走子嚢から放出される遊走子 |
|||
| image4 = Zoospores saprolegnia.svg |
|||
| caption4 = '''3d'''. ミズカビ属(ミズカビ目)の一次遊走子 (a) と二次遊走子 (b) |
|||
}} |
|||
'''[[遊走子]]は、基本的に細胞腹面(側面)から生じて前後に伸びる2本の不等[[鞭毛]]をもつ'''<ref name="Webster2007" />(上図3d(b))。例外的に、サカゲフクロカビ属({{Snamei||Anisolpidium}}; サカゲフクロカビ目)では、細胞前端付近から前鞭毛のみが生じている{{efn2|name="サカゲフクロカビ"|このため、サカゲフクロカビ属は[[サカゲツボカビ綱]]に分類されていたが、[[分子系統学]]的研究から卵菌綱に含まれることが示され、卵菌綱に移された<ref name="稲葉2013">{{cite book|author=稲葉重樹|year=2013|chapter=1.8.6 サカゲツボカビ門|editor=日本菌学会 (編)|title=菌類の事典|publisher=朝倉書店|isbn=978-4254171471|pages=41–42}}</ref><ref name="Beakes2017" /><ref name="Gachon2017">{{Cite journal|author=Gachon, C. M., Strittmatter, M., Badis, Y., Fletcher, K. I., West, P. V. & Müller, D. G.|year=2017|title=Pathogens of brown algae: culture studies of ''Anisolpidium ectocarpii'' and ''A. rosenvingei'' reveal that the Anisolpidiales are uniflagellated oomycetes|journal=European Journal of Phycology|volume=52|issue=2|pages=133-148|doi=10.1080/09670262.2016.1252857}}</ref>。}}。[[ストラメノパイル]]に属する他の生物と同様、前鞭毛には管状小毛(管状マスチゴネマ)が付随した羽型鞭毛であり、後鞭毛はこれを欠く尾型鞭毛である<ref name="Webster2007" /><ref name="Beakes2014" />(上図3d)。遊走子が着生する際には、遊走子細胞内に存在する細胞壁物質を含んだ小胞がエクソサイトーシスされて細胞壁を形成してシスト化する<ref name="Webster2007" />。 |
|||
== 分類 == |
|||
=== 基底クレード === |
|||
*{{Sname||Haptoglossales}} |
|||
**{{Snamei||Haptoglossa}} - 線虫寄生、銃細胞 (gun cell)と呼ばれる細胞から針を射出することによって寄生を行う<ref name=Beakes12>{{cite journal|author=Gordon W. Beakes & Sally L. Glockling & |
|||
Satoshi Sekimoto|title=The evolutionary phylogeny of the oomycete “fungi”|journal=Protoplasma|year=2012|volume=249|pages=3–19|doi=10.1007/s00709-011-0269-2}}</ref>。 |
|||
*{{Sname||Eurychasmales}} |
|||
**{{Snamei||Eurychasma}} - [[シオミドロ目]]の褐藻に寄生<ref name=Beakes>{{cite web|title=The evolutionary phylogeny of Oomycetes—insights gained from studies of holocarpic parasites of algae and invertebrates|url=http://media.wiley.com/product_data/excerpt/76/04702556/0470255676.pdf|accessdate=2012-10-25}}</ref> |
|||
**{{Snamei||Ectrogella}}・{{Snamei||Lagenisma}} - [[中心珪藻類]]に寄生<ref name=Beakes/> |
|||
*{{Sname||Sirolpidiaceae}} |
|||
*{{Sname||Pontismataceae}} |
|||
*{{Sname||Haliphthorales}} - 甲殻類寄生 |
|||
**{{Snamei||Haliphthoros}} |
|||
**{{Snamei||Halodaphnea}} syn. Halocrusticida |
|||
==== フクロカビモドキ目 ==== |
|||
フクロカビモドキ類 (Olpidiopsidales) は[[紅藻]]、または卵菌類の寄生者で、袋状の単細胞葉状体が葉状体の原形質内部に浸って寄生生活を送る。養殖ノリに被害を与える種が知られている。和名学名共に、[[ツボカビ]]類に属するフクロカビ(''Olpidium'')に、[[全実性]]の菌体がよく似た姿をとることによる。 |
|||
[[ファイル:Abb1.1_evolution_fungi_animals_plants_autotrophic_heterotrophic_2021_(M._Piepenbring)_198.svg|thumb|right|300px|'''4'''. ミズカビ科に見られる遊走子形成の多様性: ミズカビ属 ({{Snamei||Saprolegnia}}) は一次遊走子を放出、ヤブレワタカビ属 ({{Snamei||Thraustotheca}}) は遊走子嚢内で一次遊走子がシスト化したものが放出、アミワタカビ属 ({{Snamei||Dictyuchus}}) では遊走子嚢内で一次遊走子がシスト化したものから二次遊走子を放出、{{Snamei||Aplanes}} では遊走子嚢内で一次遊走子がシスト化したものから直接発芽、ワタカビ属 ({{Snamei||Achlya}}) では一次遊走子が遊走子嚢の開口部でシスト化し、そこから二次遊走子を放出または直接発芽する。]] |
|||
=== ミズカビ類 saprolegnian === |
|||
ミズカビ亜綱の種は、2本の[[鞭毛]]が細胞前端付近から生じている[[遊走子]]と、細胞腹面から生じている遊走子をもつことがあり、前者を一次遊走子 (primary zoospore, auxiliary zoospore)、後者を二次遊走子 (secondary zoospore, principal zoospore) とよばれる<ref name="Webster2007" /><ref name="Beakes2014" /><ref name="Beakes2017" />(上図3d, 5)。このような遊走子の二形性は二回遊泳性 (diplanetism) ともよばれる<ref name="Webster2007" /><ref name="Beakes2014" /><ref name="徳増2005" />。ミズカビ属({{Snamei||Saprolegnia}}; ミズカビ目)では、菌糸先端の遊走子嚢から放出された一次遊走子は着生、鞭毛を吸収して[[細胞壁]]を形成し[[シスト]]化し、そこから1個の二次遊走子が生じて再び遊泳する<ref name="Webster2007" /><ref name="Beakes2017" />(下図5)。二次遊走子は鞭毛を落としてシスト化し、[[菌糸]]を伸ばして発芽するか、または再び二次遊走子が形成される(数回遊泳性 polyplanetism)<ref name="Webster2007" /><ref name="Beakes2017" />。ミズカビ目の中には、これを基本形としてさまざまな様式があり、{{Snamei||Aphanomyces}} やワタカビ属 ({{Snamei||Achlya}}) では遊走子嚢の開口部で、アミワタカビ属 ({{Snamei||Dictyuchus}}) やヤブレワタカビ属 ({{Snamei||Thraustotheca}}) では遊走子嚢中で、一次遊走子に相当する細胞がシスト化し、二次遊走子を放出、または直接発芽する<ref name="Webster2007" /><ref name="Beakes2017" />(図4)。一方、{{Snamei||Phythiopsis}} は一次遊走子のみを形成し、これがシスト化して発芽する<ref name="Webster2007" />。 |
|||
最も簡単に観察できる卵菌は、[[ミズカビ]]類である。もしも[[金魚]]を飼っていれば、水底に沈んだ餌の残りや、産まれた[[卵]]が[[無精卵]]や[[発生 (生物学)|発生]]異常などで死んだときなどに、その表面に綿毛のようなものが密生するのを見る機会があるであろう。[[実験]]的には淡水中に[[スルメ]]のかけらや[[麻]]の種子をゆでたものを入れることで、たやすく観察できる。ただし、死んだものに着くものでも、時には弱った[[魚]]など生きている[[生物]]を攻撃して[[寄生]]する場合があり、[[養殖]]などに被害を与えることもある。 |
|||
[[遊走子]]は、明瞭な細胞外皮をもたない<ref name="Beakes2014" />。他の[[ストラメノパイル]]と同様、遊走子は基本的に4組の[[微小管]]性鞭毛根(R1–R4)をもつ<ref name="Beakes2014" /><ref name="Beakes2017" />。鞭毛移行部(鞭毛と基底小体の移行部)には、ふつう2重のらせん構造 (transitional helis) が存在するが、1重らせんのものやこれを欠くものもある<ref name="Beakes2014" /><ref name="Beakes2017" />。遊走子の細胞内にはさまざまなタイプの小胞が存在し、付着や外被形成に関与すると考えられている<ref name="Beakes2014" /><ref name="Beakes2017" />(上記参照)。 |
|||
ミズカビ類の遊走子嚢形成に際して基質から水中に立ち上がる菌糸は基部の直径がしばしば10 μm以上と非常に太く、肉眼でその本数が数えられるほどで、長さも1 cm程度には軽く達する。菌糸先端が区切られて遊走子嚢となり、先端に開いた穴から[[遊走子]]が泳ぎだす。遊走子嚢内の遊走子の輪郭がはっきりしてから泳ぎ出すのに数十分程度なので、[[顕微鏡]]下で泳ぎ出すのを観察するのは簡単である。 |
|||
また、卵菌では菌体の一部が厚壁化し、無性生殖の単位となることがある。この構造は菌芽 (gemma) や[[厚壁胞子]] (chlamydospore) とよばれ、休眠構造として機能し、好適な条件で発芽管を伸ばすか、または遊走子嚢を形成する<ref name="Webster2007" /><ref name="Beakes2014" /><ref name="徳増2005" />。 |
|||
{{Snamei||Aphanomyces}} 属には水生種だけではなく陸上植物寄生種が記録されている。 |
|||
=== 有性生殖 === |
|||
*{{Sname||Atkinsiellales}} |
|||
卵菌の中で生活環における核相変化が知られているものでは、'''栄養体が複相(染色体を2セットもつ)であり、配偶子形成時に減数分裂を行う'''<ref name="Beakes2017" /><ref name="徳増2005" />。ただし初期分岐群では、核相変化・有性生殖が報告されていないものが多い<ref name="Beakes2017" />。卵菌の有性生殖では、鞭毛をもつ配偶子の合体は見られず、不動状態の配偶子嚢が直接接着する('''配偶子嚢接合''' gametangial conjugation)ことで起こる<ref name="Beakes2017" />。この結果形成される接合子は'''卵胞子''' (oospore) とよばれ、ふつう休眠細胞となり、発芽して遊走子嚢を形成、または菌糸など栄養体となる<ref name="Beakes2017" /><ref name="徳増2005" />。 |
|||
**{{Snamei||Atkinsiella}} |
|||
**{{Snamei||Crypticola}} - ボウフラ寄生<ref name=Beakes/> |
|||
*[[フシミズカビ目]] {{Sname||Leptomitales}} |
|||
**Apodachlyellaceae |
|||
**Ducellieriaceae |
|||
**Leptolegniellaceae |
|||
**Leptomitaceae |
|||
*[[ミズカビ目]] {{Sname||Saprolegniales}} |
|||
**[[ミズカビ科]] {{Sname||Saprolegniaceae}} |
|||
**Leptolegniaceae |
|||
[[ファイル:Abb1.1_evolution_fungi_animals_plants_autotrophic_heterotrophic_2021_(M._Piepenbring)_200.svg|thumb|center|500px|'''5'''. ミズカビ属(ミズカビ目)の生活環: 中央の多核菌糸 (coenocytic hypha) が栄養体(通常時の体)。無性生殖(右側)では、菌糸先端が区画化されて遊走子嚢 (sporangium) となり、遊走子を放出する (二回遊泳性 diplanetism)。ミズカビ属では、放出された一次遊走子 (primary zoospore) がシスト化 (cyst) し、二次遊走子 (secondary zoospore) を放出する。有性生殖(左側)では、菌糸に生卵器 (oogonium) と造精器 (antheridium) が形成され、その中で[[減数分裂]] (M!) が起こり、生卵器と造精器が接着、細胞質合体 (P!)、核合体 (K!) が起こって受精卵(接合子)は卵胞子 (oospore) になる。]] |
|||
=== フハイカビ類 peronosporalean === |
|||
[[ファイル:07 08 life cycle, Phytophthora infestans on potato, Peronosporales, Oomycota (M. Piepenbring).png|thumb|center|500px|'''6'''. [[ジャガイモエキビョウキン]](ツユカビ目)の生活環: [[ジャガイモ]]に感染 (infection) して栄養体(通常時の体)である[[菌糸]]を葉や地下茎内に伸ばす (中央)。[[無性生殖]](asexual; 右側)では、葉裏の[[気孔]]から伸ばした分枝した胞子嚢柄に胞子嚢 (sporangium) をつけ、そこから遊走子 (zoospore) を放出、または胞子嚢 ([[分生子]] conidium ともよばれる) が散布されて直接発芽する。有性生殖(sexual; 左側)では、菌糸に造精器 (antheridium) とそれを貫く生卵器 (oogonium) が形成され、その中で[[減数分裂]] (M!) が起こり、細胞質合体 (P!)、核合体 (K!) が起こって受精卵(接合子)は卵胞子 (oospore) になる。]] |
|||
[[ツユカビ目]]の多くは陸上植物の地上部に[[寄生]]する[[絶対寄生菌]]で、植物に[[べと病]]を引き起こすツユカビ科、[[白さび病]]を引き起こすシロサビキン科などが知られる。ツユカビ類は、[[接合菌門]]の[[ケカビ]]類に見られるような、宿主から立ち上がった胞子嚢柄の先端に胞子嚢を形成する。この胞子嚢自体が[[カビ]]の胞子のように飛散して宿主に到達する。この胞子嚢はミズカビの遊走子嚢と相同であり、宿主に到達し、そこに水があると[[遊走子]]を放出する。ツユカビ目には二次型遊走子しか見られない。遊走子を放出せず、直接葉状体が発芽するものもあり、いずれにせよ宿主の組織内に侵入し、宿主の細胞内に[[吸器]]が差し込まれ、寄生が成立する。 |
|||
{{Snamei||Eurychasma}}(ユーリカスマ目)や {{Snamei||Anisolpidium}}(サカゲフクロカビ目)、{{Snamei||Lagenisma}}(フシミズカビ目)では、鞭毛細胞が着生してシスト化したものが受精管 (fertilization tube) でつながり、一方の原形質がもう一方へ送り込まれ、細胞質・核融合が起きて接合子が形成される<ref name="Beakes2017" />。{{Snamei||Lagenisma coscinodisci}}では、減数分裂によって形成された遊走子が着生して接合することが報告されている<ref name="Schnepf1977">{{Cite journal|author=Schnepf, E. & Drebes, G.|year=1977|title=On the development of the marine, parasitic phycomycete ''Lagenisma coscinodisci'' (Lagenidiales)|journal=Helgoländer Wissenschaftliche Meeresuntersuchungen|volume=29|issue=|pages=291-301|doi=10.1007/BF01614265}}</ref>。このような雌雄の分化がない同型の配偶子嚢接合が卵菌における原始形質であると考えられている<ref name="Beakes2017" />。 |
|||
[[フハイカビ科]]にも水生種に加えて陸上植物寄生種が数多く知られ、[[19世紀]]に[[ジャガイモ]]に壊滅的な被害を与えて[[アイルランド]]に大[[飢饉]]([[ジャガイモ飢饉]])をもたらした[[疫病]]の病原体も卵菌のフハイカビ科に属する[[エキビョウキン]] (''Phytophthora'') である。フハイカビ科では他に[[フハイカビ]](''Pythium'')、''Phytopythium''、''Sclerophthora''に陸上植物に寄生する種がある。 |
|||
ミズカビ亜綱やツユカビ亜綱の多くでは、配偶子嚢の雌雄の分化が見られる<ref name="Webster2007" /><ref name="Beakes2017" />。雌雄配偶子嚢内では、[[減数分裂]]が起こり、[[核相|単相]]([[染色体]]を1セットのみもつ)の核を形成する<ref name="Webster2007" /><ref name="Beakes2017" />。雌性配偶子嚢である'''生卵器''' (oogonium) は、1個から数個の単相の[[卵細胞|卵]](卵球 oosphere ともよばれる)を形成する<ref name="Webster2007" /><ref name="Beakes2017" /><ref name="徳増2005" />。雄性配偶子嚢である'''造精器''' (antheridium) は生卵器に接し(配偶子嚢接着 gametangial contact)、受精管を通じて単相の雄性核を送り込み、卵において受精が起こる<ref name="Webster2007" /><ref name="Beakes2017" /><ref name="徳増2005" />。配偶子嚢や卵胞子の形態は、このような卵菌の分類において極めて重視されている<ref name="Beakes2017" />。このような有性生殖は、[[ステロイド]]性の[[ホルモン]]によって制御されていることが知られている<ref name="Webster2007" /><ref name="Beakes2017" />。ツユカビ亜綱は[[ステロール]]合成能を欠き、有性生殖のためには外部からステロールを得る必要がある<ref name="Webster2007" /><ref name="Beakes2017" />。 |
|||
*[[オオギミズカビ目]] {{Sname||Rhipidiales}} - 腐生性、淡水中の果実など |
|||
**[[オオギミズカビ科]] {{Sname||Rhipidiaceae}} |
|||
有性生殖は、[[ホモタリック]](雌雄同株)のものと、[[ヘテロタリック]](雌雄異株)のものが知られている<ref name="Beakes2017" />。ホモタリックの場合、生卵器と造精器のできる位置関係には多様性があり、造精器が生卵器の直下にできるもの (直下性 hypogynous)、生卵器と同一の柄に側生するもの (雌雄同菌糸性 monoclinous)、同一の菌体の異なる菌糸にできるもの (雌雄異菌糸性 diclinous) がある<ref name="Beakes2017" /><ref name="東條2004">{{Cite journal|author=東條元昭|year=2004|title=ピシウム菌による病害|journal=植物防疫|volume=58|issue=3|pages=120–126|url=http://jppa.or.jp/archive/pdf/58_03_24.pdf}}</ref>。[[エキビョウキン|エキビョウキン属]](ツユカビ目)では、生卵器が造精器を貫通して発達し、生卵器の基部を造精器が取り巻いている状態 (amphigynous) になるものが多い<ref name="Beakes2017" />(上図6)。また、ヘテロタリックの場合、相手によって雌雄性が切り替わる相対的雌雄性 (relative sexuality) を示すものも知られており、相手が生卵器を形成する場合は造精器を、相手が造精器を形成する場合は生卵器を形成する<ref name="Webster2007" />。また一部の種では、造精器と接着せずに、1個の生卵器の中で2個の単相核が合体することや、減数分裂せずに複相の核をもとに卵胞子が形成されることがある<ref name="Webster2007" />。 |
|||
*[[シロサビキン目]] {{Sname||Albuginales}} |
|||
**シロサビキン科 Albuginaceae |
|||
生卵器内での卵形成様式には多様性があり、ミズカビ亜綱では遠心的に、ツユカビ亜綱では求心的に卵が形成される<ref name="Beakes2017" />。またツユカビ亜綱では、周辺の細胞質が卵にならずに取り残されて周辺細胞質 (periplasm) となり、卵胞子の細胞壁形成などに寄与する<ref name="Beakes2017" />。 |
|||
*[[ツユカビ目]] {{Sname||Peronosporales}} |
|||
**ツユカビ科 Peronosporaceae |
|||
卵胞子内にはooplastとよばれる大きな[[液胞]]([[マイコラミナリン]]や[[リン酸]]を含む)が存在し、また周縁部には脂質顆粒(油滴)が存在する<ref name="Webster2007" /><ref name="Beakes2017" />。このooplastと油滴の配置様式に基づいて下記のようにいくつかの型に分けられており、分類形質として用いられている<ref name="Webster2007" /><ref name="Beakes2017" /><ref name="印東1985">{{Cite journal|author=印東弘玄|year=1985|title=水棲菌類の観察方法|journal=日本水処理生物学会誌|volume=21|issue=1|pages=20-25|doi=10.2521/jswtb.21.20}}</ref>。 |
|||
*[[フハイカビ目]] {{Sname||Pythiales}} |
|||
**[[フハイカビ科]] {{Sname||Pythiaceae}} |
|||
* セントリック (中心生、centric) … ooplastが中央に位置しており、その周囲を1–2層の小脂質顆粒が囲んでいる。 |
|||
***[[クサリフクロカビ属]] {{Snamei||Lagenidium}} |
|||
* サブセントリック (亜中心生、subcentric) … ooplastがやや偏って位置しており、周辺の小脂質顆粒が多層の部分と1–0層の部分がある。 |
|||
*{{Sname||Myzocytiopsidales}} - 線虫寄生 |
|||
* サブエクセントリック (subeccentric) … ooplastが偏って位置しており、その反対側に複数の脂質顆粒がある。 |
|||
**Myzocytiopsidaceae |
|||
* エクセントリック (偏心生、eccentric) … ooplastが偏って位置しており、その反対側に1個の大きな脂質顆粒がある。 |
|||
*{{Sname||Salilagenidiales}} - 海棲 |
|||
卵胞子は厚くふつう多層の[[細胞壁]]に囲まれており、また突起などで装飾されていることもある<ref name="Beakes2017" />。卵胞子の細胞壁内層はendospore、外層はepisporeとよばれる<ref name="Webster2007" />。卵胞子は耐久構造となり、ふつう休眠ののちに発芽して発芽管を伸ばす。ふつう発芽管の先端に遊走子嚢を形成するが、直接菌糸を伸ばすことや、卵胞子内で遊走子が形成されることもある<ref name="Beakes2017" />。 |
|||
== 生態 == |
|||
卵菌は海や淡水、陸上に広く分布する<ref name="Webster2007" /><ref name="Beakes2017" />。栄養様式としては、腐生性のものと寄生性のものがいる。 |
|||
腐生性の種は、植物遺体や動物遺体を分解する<ref name="Beakes2017" /><ref name="徳増2005" />(下図7a, b)。淡水域には、ミズカビ目の腐生性の種が極めてふつうに見られる<ref name="Beakes2017" />。一般的に、このような卵菌は湖沼の沿岸域に多く、四季がある地域では春と秋にピークがある<ref name="Beakes2017" />。また海の沿岸域でも、{{Snamei||Halophytophthora}} や {{Snamei||Salisapilia}}(ツユカビ目)が、[[マングローブ林]]や[[塩性湿地]]の植物質の初期分解に重要な役割を担っている<ref name="Beakes2017" />。 |
|||
カップに水サンプルまたは水と土壌サンプルや水中の枯れ枝などを入れ、これに基質となる[[アサ]]の実や[[ハエ]]の死骸などを加えておくことによって、数日で腐生性の卵菌、特にミズカビ科の菌糸が見られるようになる(このような手法は釣菌法(ちょうきんほう)とよばれる)<ref name="Webster2007" /><ref name="稲葉2014">{{Cite book|author=稲葉重樹|year=2014|chapter=鞭毛菌類|editor=細矢剛, 国立科学博物館|title=菌類のふしぎ 第2版|publisher=東海大学出版部|isbn=978-4486020264|pages=20–28}}</ref>。 |
|||
卵菌の多くは[[好気性]]であるが、フシミズカビ目やオオギミズカビ目、ツユカビ目の中には、嫌気的水域で[[発酵|有機酸発酵]]などによって生育する[[通性嫌気性生物|通性嫌気性]]のものも知られている<ref name="Webster2007" /><ref name="Beakes2017" />。 |
|||
寄生性の種の中には、宿主の一部を殺して分解・吸収する[[殺生栄養]]性のものと、宿主を生かした状態で栄養を吸収する[[活物栄養]]性のものがある<ref name="Webster2007" /><ref name="Beakes2014" /><ref name="Beakes2017" /><ref name="徳増2005" />。腐生性と殺生栄養性の間は連続的であり、条件によって変わるものもいる<ref name="Webster2007" /><ref name="Beakes2014" />。このような条件的寄生性卵菌の中には、{{Snamei||Aphanomyces}}(ミズカビ目)、[[フハイカビ|フハイカビ属]]({{Snamei||Pythium}})、[[エキビョウキン|エキビョウキン属]]({{Snamei||Phytophthora}})(ツユカビ目)など土壌生態系において分解者としても重要な存在もいる<ref name="Beakes2017" />。活物栄養性の種は、ふつう絶対寄生性(寄生しなければ生きられない)である<ref name="Webster2007" />。 |
|||
寄生性の種は、[[陸上植物]]に寄生するものが多く知られており、組織の細胞間に[[菌糸]]を伸ばしている<ref name="Beakes2017" /><ref name="徳増2005" />(上図2c, 下図7c, d)。宿主はふつう草本性の被子植物であるが、木本に寄生する種もいる<ref name="Beakes2017" />。また、宿主特異性が比較的高いものが多いが、様々な植物に寄生する宿主範囲が広いもの(ツユカビ属など)もいる<ref name="Beakes2017" />。 |
|||
{{multiple image |
|||
| total_width = 800 |
|||
| align = center |
|||
| caption_align = left |
|||
| image1 = Water mold Mizukabi colony.jpg |
|||
| caption1 = '''7a'''. [[アサ]]の実から生じたミズカビ類 |
|||
| image2 = Water mold.JPG |
|||
| caption2 = '''7b'''. 昆虫の遺骸から生じたミズカビ類 |
|||
| image3 = Albugo candida on Capsella bursa-pastoris kz01.jpg |
|||
| caption3 = '''7c'''. 白さび病の[[ナズナ]] |
|||
| image4 = Falscher Mehltau.jpg |
|||
| caption4 = '''7d'''. キュウリべと病 |
|||
}} |
|||
陸上植物以外にも、他の卵菌や[[菌類]]、[[海藻]]([[紅藻]]、[[緑藻]]、[[褐藻]])、微細藻([[珪藻]]、[[接合藻]]など)に寄生する卵菌も多く知られている<ref name="Beakes2017" /><ref name="Buaya2019" /><ref name="Buaya2021a" /><ref name="Buaya2021b" /><ref name="Buaya2020" /><ref name="Buaya2019a">{{Cite journal|author=Buaya, A. T., Ploch, S., & Thines, M.|year=2019|title=Rediscovery and phylogenetic placement of ''Olpidiopsis gillii'' (de Wildeman) Friedmann, a holocarpic oomycete parasitoid of freshwater diatoms|journal=Mycoscience|volume=60|issue=3|pages=141-146|doi=}}</ref><ref name="Buaya2020b">{{Cite journal|author=Buaya, A. T., & Thines, M.|year=2020|title=Diatomophthoraceae–a new family of olpidiopsis-like diatom parasitoids largely unrelated to ''Ectrogella''|journal= Fungal Systematics and Evolution|volume=5|issue=1|pages=113-118|doi=}}</ref><ref name="Buaya2020c">{{Cite journal|author=Buaya, A. T. & Thines, M.|year=2020|title=''Bolbea parasitica'' gen. et sp. nov., a cultivable holocarpic parasitoid of the early-diverging Saprolegniomycetes|journal=Fungal Systematics and Evolution|volume=6|issue=1|pages=129-137|doi=}}</ref><ref>{{Cite journal|author=竹中重仁|year=2011|title=菌寄生菌としての ''Pythium''|journal=植物防疫|volume=65|issue=2|pages=117-120|crid=1522543655605926784}}</ref><ref name="築尾2013">{{Cite journal|author=築尾嘉章 & 東條元昭|year=2013|title=疫病菌とピシウム菌 類似点と相違点|journal=植物防疫|volume=67|issue=10|pages=536-552|crid=1521136279780135808}}</ref>。これらの多くは全実性(細胞全体が遊走子嚢になる)であり、宿主細胞内で生育する。 |
|||
卵菌の中には動物に寄生するものもおり、宿主として[[甲殻類]]、[[昆虫]]、[[線虫]]、[[ワムシ]]などが知られている<ref name="Webster2007" /><ref name="Beakes2017" />。{{Snamei||Haptoglossa}}(ハプトグロッサ目)では、[[遊走子]]がシスト化し、銃細胞 (gun cell) とよばれる特殊な細胞となり、線虫に触れると銃弾状の構造を撃ち込み、これを通って原形質が線虫内に侵入する<ref name="Beakes2017" />。 |
|||
== 人間との関わり == |
|||
植物寄生性の卵菌の中には農作物など有用植物に寄生して被害を与える種が多く(下図8)、代表的なものを下表1に示した。[[ジャガイモエキビョウキン]]({{Snamei||Phytophthora infestans}}; [[ツユカビ目]])は[[ジャガイモ]]に寄生し、大きな被害を与えることがある(下図8a)。特に19世紀半ば(1845–1847)に[[ジャガイモ]]を主食としていた[[アイルランド]]で大飢饉([[ジャガイモ飢饉]])を引き起こし、大量の餓死者と北米への移民を生み出したため、1841年から1851年の間にアイルランドの人口は162万人も減少した<ref name="ウェブスター">{{Cite book|author=ジョン・ウェブスター著|translator=椿啓介、三浦宏一郎、山本昌木|year=1985|chapter=卵菌綱|editor=|title=ウェブスター菌類概論|publisher=講談社|isbn=978-4061396098|pages=139–187}}</ref>。また、19世紀のヨーロッパでは、北米から侵入したブドウべと病菌({{Snamei||Plasmopara viticola}}; ツユカビ目)によってブドウ栽培に大きな被害が生じた<ref name="Fontaine2021">{{Cite journal|author=Fontaine, M. C., Labbé, F., Dussert, Y., Delière, L., Richart-Cervera, S., Giraud, T. & Delmotte, F.|year=2021|title=Europe as a bridgehead in the worldwide invasion history of grapevine downy mildew, ''Plasmopara viticola''|journal=Current Biology|volume=31|issue=10|pages=2155-2166|doi=10.1016/j.cub.2021.03.009 }}</ref>(下図8b)。その対策として、[[硫酸銅]]、[[生石灰]]、水の混合液である[[ボルドー液]]が使われた<ref name="Webster2007" />。1990年代には、米国[[カリフォルニア州]]のマテバシイ属やコナラ属の森林において {{Snamei||Phytophthora ramorum}} によるブナ科樹木の急激な枯死 (オーク突然死 Sudden Oak Death) が起こり、大きな被害が生じた<ref name="Rizzo2003">{{Cite journal|author=Rizzo, D. M. & Garbelotto, M.|year=2003|title=Sudden oak death: endangering California and Oregon forest ecosystems|journal=Frontiers in Ecology and the Environment|volume=1|issue=4|pages=197-204|doi=10.1890/1540-9295(2003)001[0197:SODECA]2.0.CO;2}}</ref><ref name="升屋2019">{{Cite journal|author=升屋勇人, 田端雅進, 市原優 & 景山幸二|year=2019|title=''Phytophthora cinnamomi'' によるウルシ林の衰退 ―国産漆の新たなる脅威―|journal=日本森林学会誌|volume=101|issue=6|pages=318-321|doi=10.4005/jjfs.101.318}}</ref>(下図8c)。また、[[オーストラリア]]には {{Snamei||Phytophthora cinnamomi}} が侵入し、森林生態系に多大な損害を与えている<ref name="升屋2019" />。 |
|||
{{multiple image |
|||
| total_width = 800 |
|||
| align = center |
|||
| caption_align = left |
|||
| image1 = Aardappel Phytophthora Fresco.jpg |
|||
| caption1 = '''8a'''. ジャガイモ疫病菌に寄生されたジャガイモ |
|||
| image2 = Plasmopara viticola a1 (4).jpg |
|||
| caption2 = '''8b'''. ブドウべと病 |
|||
| image3 = Dead Coast Live Oak in Marin.Steve Swain-1- (5812704230).jpg |
|||
| caption3 = '''8c'''. {{Snamei||Phytophthora ramorum}} によって枯れたナラ (Sudden Oak Death) |
|||
| image4 = Typical symptom of maize downy mildew (Peronosclerospora maydis) Pj DSC 0774.jpg |
|||
| caption4 = '''8d'''. トウモロコシべと病 |
|||
}} |
|||
{| class="wikitable" style="margin:0 auto;" |
|||
|+ 表1. <span style="font-weight:400;">植物病害性の卵菌<ref name="埋橋2011">{{Cite journal|author=埋橋志穂美|year=2011|title=分子系統に基づく ''Pythium'' 属の新分類システム|journal=植物防疫|volume=65|issue=10|pages=587-592|crid=1522543655625413888}}</ref><ref name="日本植物病名データベース">{{Cite web|author=|date=|url=https://www.gene.affrc.go.jp/databases-micro_pl_diseases.php|title=日本植物病名データベース|website=|publisher=農業生物資源ジーンバンク|accessdate=2023-09-12}}</ref><ref name="植物病原菌類図説1992">{{cite book|author=|year=1992|chapter=|editor=小林亨夫・勝本謙・安孫子和雄・阿部恭久・柿島真 (編)|title=植物病原菌類図説|publisher=全国農村教育協会|isbn=9784881370452|pages=58–71, 510–512}}</ref></span> |
|||
! 目 !! 科 !! 属 !! 病例 |
|||
|- |
|||
| ミズカビ目 || ベルカルブス科 || {{Snamei||Aphanomyces}} || ハクサイ根くびれ病、テンサイ黒根病、ホウレンソウ根腐病など |
|||
|- |
|||
| シロサビキン目 || シロサビキン科 || シロサビキン属 {{Snamei||Albugo}} || ダイコン白さび病、ハクサイ白さび病、ワサビ白さび病、サツマイモ白さび病など |
|||
|- |
|||
| rowspan="13" | ツユカビ目 || rowspan="2" | フハイカビ科 || [[フハイカビ|フハイカビ属]] {{Snamei||Pythium}} || イネ苗立枯病、トウモロコシ根腐病、ムギ類褐色雪腐病、シバピシウム病、チューリップピシウム葉枯病、ホウレンソウ立枯病、キャベツピシウム腐敗病、スイカ綿腐病、トマト根腐病、サツマイモ白腐病、ニンジンしみ腐病など |
|||
|- |
|||
| {{Snamei||Globisporangium}} || イネ苗立枯病、ムギ類褐色雪腐病、ホウレンソウ立枯病、テンサイ苗立枯病、キャベツ苗立枯病、ブロッコリーピシウム腐敗病、レタスピシウム萎凋病など |
|||
|- |
|||
| rowspan="11" | ツユカビ科 || {{Snamei||Phytopythium}} || ブロッコリーピシウム腐敗病、イチゴピシウム根腐病、キウイフルーツ根腐病、トマト苗立枯病、バジル根腐病、キクピシウム立枯病など |
|||
|- |
|||
| [[エキビョウキン|エキビョウキン属]] {{Snamei||Phytophthora}} || ジャガイモ疫病、キュウリ疫病、レタス疫病、イチゴ疫病、ナシ疫病、ネギ白色疫病、アスパラガス疫病、パイナップル心腐病など |
|||
|- |
|||
| {{Snamei||Sclerophthora}} || イネ黄化萎縮病、コムギ黄化萎縮病、トウモロコシ褐条べと病など |
|||
|- |
|||
| ササラビョウキン属 {{Snamei||Sclerospora}} || アワしらが病など |
|||
|- |
|||
| タンジクツユカビ属 {{Snamei||Plasmopara}} || ブドウべと病、ミツバべと病、ヒマワリべと病など |
|||
|- |
|||
| ラッパツユカビ属 {{Snamei||Bremia}} || シュンギクべと病、レタスべと病など |
|||
|- |
|||
| {{Snamei||Hyaloperonospora}} || ハクサイべと病、キャベツべと病など |
|||
|- |
|||
| {{Snamei||Peronosclerospora}} || トウモロコシべと病、サトウキビしらが病など |
|||
|- |
|||
| ツユカビ属 {{Snamei||Peronospora}} || タマネギべと病、ホウレンソウべと病、ダイズべと病、ソバべと病、タバコべと病、ブロッコリーべと病、バジルべと病など |
|||
|- |
|||
| ニセツユカビ属 {{Snamei||Pseudoperonospora}} || キュウリべと病、ホップべと病、タイマべと病、エノキべと病など |
|||
|} |
|||
養殖海藻である[[アマノリ]]([[紅藻]])に寄生して被害を与える卵菌がいくつか知られており、{{Snamei||Pontisma porphyrae}} (= {{Snamei|Olpidiopsis porphyrae}}) は壺状菌病を<ref name="Sekimoto2008">{{Cite journal|author=Sekimoto, S., Yokoo, K., Kawamura, Y. & Honda, D.|year=2008|title=Taxonomy, molecular phylogeny, and ultrastructural morphology of ''Olpidiopsis porphyrae'' sp. nov.(Oomycetes, straminipiles), a unicellular obligate endoparasite of ''Bangia'' and ''Porphyra'' spp.(Bangiales, Rhodophyta)|journal=Mycological Research|volume=112|issue=3|pages=361-374|doi=10.1016/j.mycres.2007.11.002}}</ref><ref name="Buaya2019">{{Cite journal|author=Buaya, A. T., Ploch, S., Inaba, S., & Thines, M.|year=2019|title=Holocarpic oomycete parasitoids of red algae are not ''Olpidiopsis''|journal=Fungal Systematics and Evolution|volume=4|issue=1|pages=21-31|doi=10.3114/fuse.2019.04.03}}</ref>、{{Snamei||Pythium porphyrae}} はあかぐされ病を<ref>{{Cite journal|author=東條元昭|year=2009|title=ノリアカグサレ菌の分類学的研究|journal=日本水産学会誌|volume=75|issue=5|pages=910-911|doi=10.2331/suisan.75.910}}</ref>、それぞれ引き起こす。 |
|||
[[ミズカビ属]]やワタカビ属、{{Snamei||Aphanomyces}}([[ミズカビ目]])などの卵菌は、淡水養殖魚(特に[[サケ]])に寄生して大きな被害を与えることがある<ref name="Webster2007" />。また、{{Snamei||Pythium flevoense}}(ツユカビ目)は、[[アユ]]仔魚の内臓真菌症を引き起こすことが報告されている<ref>{{Cite journal|author=三浦正之, 畑井喜司雄, 東條元昭, 和田新平, 小林咲麗 & 岡崎巧|year=2010|title=アユ仔魚に発生した ''Pythium flevoense'' による内臓真菌症|journal=魚病研究|volume=45|issue=1|pages=24-30|doi=10.3147/jsfp.45.24}}</ref>。 |
|||
[[ザリガニ]]に寄生する[[北米]]産の {{Snamei||Aphanomyces astaci}}([[ミズカビ目]])は抵抗性をもたない他地域のザリガニに水カビ症(ザリガニペストともよばれる)を引き起こし(下図9a)、[[スウェーデン]]では在来ザリガニ個体群の99%が消失したともされる<ref name="上村2018">{{Cite journal|author=上村兼輔 & 川井唯史|year=2018|title=新しい脅威である水カビ病について|journal=Cancer|volume=27|issue=|pages=153-154|doi=10.18988/cancer.27.0_153}}</ref>。そのため、この卵菌は[[国際自然保護連合]](IUCN)が定める「[[世界の侵略的外来種ワースト100]]」にも選定されている<ref name="上村2018" />。この卵菌は日本でも確認されており、在来ニホンザリガニへの影響が懸念されている<ref name="上村2018" />。 |
|||
{{multiple image |
|||
| total_width = 400 |
|||
| align = center |
|||
| caption_align = left |
|||
| image1 = Aphanomyces astaci colonization and immune reaction in North American crayfish - Pone.0195353.g002.png |
|||
| caption1 = '''9a'''. ザリガニの水カビ症 |
|||
| image2 = Pythiosis 2.jpg |
|||
| caption2 = '''9b'''. イヌのピシウム症 |
|||
}} |
|||
[[フハイカビ|フハイカビ属]]の {{Snamei||Pythium insidiosum}}([[ツユカビ目]])は、ヒトを含む[[哺乳類]]に対する[[日和見感染|日和見的な寄生菌]]となり、ピシウム症 (pythiosis) とよばれる皮膚病を引き起こす<ref name="稲葉2014" /><ref name="Gaastra2010">{{Cite journal|author=Gaastra, W., Lipman, L. J., De Cock, A. W., Exel, T. K., Pegge, R. B., Scheurwater, J., ... & Mendoza, L.|year=2010|title=''Pythium insidiosum'': an overview|journal=Veterinary Microbiology|volume=146|issue=1-2|pages=1-16|doi=10.1016/j.vetmic.2010.07.019}}</ref>(上図9b)。また、{{Snamei||Paralagenidium karlingii}}(ツユカビ目)も、[[イヌ]]に寄生して皮膚炎 (paralagenidiosis) を引き起こす<ref name="Beakes2017" />。 |
|||
== 系統と分類 == |
|||
=== 上位分類 === |
|||
卵菌は[[吸収栄養]]性の[[従属栄養]]生物であり、またその形態や生活様式は[[菌類]](真菌)とよく似ている。そのため、20世紀後期までは、卵菌は菌類に含められ、菌類の中では、[[ツボカビ門|ツボカビ類]]などとともに[[藻菌類]](子嚢菌・担子菌以外の真菌)や鞭毛菌類(鞭毛細胞をもつ真菌)に分類されることが多かった<ref name="徳増2005" /><ref name="ウェブスター" /><ref name="井上1983">{{cite book|author=井上浩, 岩槻邦男, 柏谷博之, 田村道夫, 堀田満, 三浦宏一郎 & 山岸高旺|year=1983|chapter=|editor=|title=植物系統分類の基礎|publisher=北隆館|isbn=|page=23–34}}</ref><ref name="杉山2005">{{cite book|author=杉山純多|year=2005|chapter=菌類の多様性と分類体系|editor=杉山純多|title=バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統|publisher=裳華房|isbn=978-4785358273|pages=30–55}}</ref>。しかし、鞭毛細胞の形態や細胞壁組成、リシン合成経路などさまざまな点で、卵菌類は狭義の菌類と異なることが認識されるようになり(下表2)、20世紀末ごろには菌類とは系統的に全く異なる生物群であると考えられるようになった<ref name="Webster2007" /><ref name="徳増2005" /><ref name="杉山2005" /><ref name="稲葉2014解体" />。卵菌の鞭毛細胞や一部の生化学的な特徴は、[[不等毛藻]]([[褐藻]]や[[珪藻]])や[[ラビリンチュラ類]]、一部の[[原生動物]]と共通しており、これらが単系統群を構成していると考えられるようになり、この生物群に対して'''[[ストラメノパイル]]''' (stramenopiles; 不等毛類) の名が提唱された<ref name="Patterson1989">{{cite book|author=Patterson, D.J.|year=1989|chapter=Stramenopiles: chromophytes from a protistan perspective|editor=Green, J.C., Leadbeater, B.S.C. & Diver, W.L.|title=Chromophyte Algae: Problems and Perspectives|publisher=Clarendon Press|isbn=0198577133|pages=357–379}}</ref>。その直後ころから[[分子系統学]]的研究が一般的となり、卵菌とこれらの生物との近縁性、ストラメノパイルの存在が支持された<ref name="Webster2007" /><ref name="杉山2005" /><ref name="稲葉2014解体" />。2023年現在では、卵菌はストラメノパイルの一群として認識されている。卵菌と菌類の間に見られるさまざまな類似点([[菌糸]]など)は、類似した生き方をするため起こった収斂進化の結果であると考えられている<ref name="Webster2007" />。 |
|||
{| class="wikitable" style="margin:0 auto;" |
|||
|+ 表2. <span style="font-weight:400;">ツボカビ類と卵菌類の比較<ref name="徳増2005" /><ref name="稲葉2014解体">{{Cite book|author=稲葉重樹|year=2014|chapter=除外された偽菌類|editor=細矢剛, 国立科学博物館|title=菌類のふしぎ 第2版|publisher=東海大学出版部|isbn=978-4486020264|pages=99–105}}</ref><ref name="Spring2012" /></span> |
|||
! !! [[ツボカビ門|ツボカビ類]]([[菌類]]) !! 卵菌類 |
|||
|- |
|||
! [[細胞壁]]主要多糖 |
|||
| [[キチン]] || [[セルロース]] |
|||
|- |
|||
! 転流炭水化物 |
|||
| [[ポリオール]]、[[トレハロース]] || [[グルコース]] |
|||
|- |
|||
! 貯蔵多糖 |
|||
| [[グリコーゲン]] || [[マイコラミナリン]] |
|||
|- |
|||
! [[リシン]]合成 |
|||
| AAA経路 || DAP経路 |
|||
|- |
|||
! [[ミトコンドリア]]クリステ |
|||
| 板状 || 管状 |
|||
|- |
|||
! ミトコンドリアの[[コドン]] |
|||
| UGAは[[トリプトファン]] || UGAは終止コドン |
|||
|- |
|||
! [[ステロール]] |
|||
| [[エルゴステロール]] || [[フコステロール]] |
|||
|- |
|||
! [[ニコチン酸]]合成 |
|||
| トリプトファンから || C3前駆体から |
|||
|- |
|||
! [[微小管]] |
|||
| [[ベンゾイミダゾール]]および<br />[[グリセオフルビン]]感受性 || [[コルヒチン]]感受性 |
|||
|- |
|||
! [[鞭毛]] |
|||
| 細胞後端から後方へ尾型鞭毛 || 細胞前端または側面から<br />前方へ羽型鞭毛、後方へ尾型鞭毛 |
|||
|} |
|||
[[分子系統学]]的研究から、ストラメノパイルの中で、卵菌は[[サカゲツボカビ類]]の[[姉妹群]]であることが示唆されている<ref name="Leonard2018">{{Cite journal|author=Leonard, G., Labarre, A., Milner, D. S., Monier, A., Soanes, D., Wideman, J. G., ... & Richards, T. A.|year=2018|title=Comparative genomic analysis of the ‘pseudofungus’ Hyphochytrium catenoides|journal=Open Biology|volume=8|issue=1|pages=170184|doi=10.1098/rsob.170184}}</ref><ref name="Cho2022">{{Cite journal|author=Cho, A., Tikhonenkov, D. V., Hehenberger, E., Karnkowska, A., Mylnikov, A. P. & Keeling, P. J.|year=2022|title=Monophyly of diverse Bigyromonadea and their impact on phylogenomic relationships within stramenopiles|journal=Molecular Phylogenetics and Evolution|volume=171|issue=|pages=107468|doi=10.1016/j.ympev.2022.107468}}</ref>。また、[[珪藻]]細胞内に侵入する捕食者である[[ピルソニア類]]や、自由生活性[[鞭毛虫]]である[[デヴェロパイエラ類]]が、卵菌類+[[サカゲツボカビ類]]に近縁であることが示唆されている<ref name="Cho2022" />。分類学的には、しばしばこれらを合わせて[[偽菌類|偽菌門]] (Pseudofungi) に分類する<ref name="Cho2022" /><ref name="辞典2013分類表" />。ストラメノパイル内において、偽菌門の姉妹群は、[[不等毛藻]](オクロ植物門)であると考えられている<ref name="Cho2022" />。ストラメノパイルの中で菌類的な生物群として、他に[[ラビリンチュラ類]]があるが(ラビリンチュラ類のうちヤブレツボカビ類とよばれる一群は、古くは卵菌に分類されていた<ref name="井上1983" />)、ストラメノパイルの中でラビリンチュラ類はより初期に分岐しており、卵菌類と近縁ではない<ref name="Beakes2017" />。ストラメノパイルの共通祖先が[[紅藻]]との二次共生由来の葉緑体をもっており、偽菌類やラビリンチュラ類などの祖先で二次的に葉緑体・光合成能を失ったとする仮説([[クロムアルベオラータ仮説]])もあるが<ref>{{Cite journal|author=矢吹彬憲・松本拓也|year=2007|title=クリプト藻類・ハプト藻類聞の姉妹群関係とクロムアルベオラータ仮説|journal=藻類|volume=55|issue=|pages=192-194|doi=}}</ref>、これを積極的に支持する証拠は見つかっていない<ref>{{Cite journal|author=Archibald, J. M.|year=2009|title=The puzzle of plastid evolution|journal=Current Biology|volume=19|issue=2|pages=R81-R88|doi=10.1016/j.cub.2008.11.067}}</ref>。 |
|||
=== 下位分類 === |
|||
卵菌は比較的大きなグループであり、2020年現在でおよそ100属1,700種ほどが知られている<ref name="Buaya2020" />。卵菌類は比較的古くから1つの綱、'''卵菌綱''' ({{Sname||Oomycetes}}) にまとめられていた<ref name="ウェブスター" /><ref name="井上1983" />。また、同じ範囲の生物群に対して、ツユカビ綱 ({{Sname|Peronosporomycetes}}) の名も提唱されている<ref name="Beakes2014" />。ただし、卵菌を1つの門、'''卵菌門''' ({{Sname||Oomycota}}) として扱い、その一部のみに対してツユカビ綱の名を用いることもある<ref name="Beakes2014" /><ref name="Beakes2017" />(下記参照)。 |
|||
菌体の体制や無性生殖様式、有性生殖様式、さらに20世紀末以降は分子形質に基づいて分類され、2023年現在では12目程度に分けられることが多い<ref name="Beakes2017" /><ref name="Buaya2020" />(下表3)。系統的には、これらの目は、多数の初期分岐群と2つの大きなグループ、ミズカビ群とツユカビ群に分けられる<ref name="Beakes2014" /><ref name="Beakes2017" />(下図10)。初期分岐群(basal oomycetes)はユーリカスマ目やフクロカビモドキ目など全て微細な内部寄生性種であり、特に2020年ごろから詳細に研究され多数知られるようになった<ref name="Beakes2017" />。ミズカビ群はフシミズカビ目(ときにアトキンシエラ目を分ける)とミズカビ目を、ツユカビ群はオオギミズカビ目、シロサビキン目、ツユカビ目を含む<ref name="Beakes2017" /><ref name="Buaya2020" />。この2つの群は亜綱(ミズカビ亜綱、ツユカビ亜綱)に分類されることもあるが、卵菌類を門とし、この2群を綱(ミズカビ綱、ツユカビ綱(狭義))として扱うこともある<ref name="Beakes2014" /><ref name="Beakes2017" />。ただし、これらの場合の初期分岐群の分類学的扱い(おそらく複数の亜綱または綱に分割する必要がある)は提唱されていない。ミズカビ群とツユカビ群は単系統群を形成すると考えられており、この単系統群は core oomycetes とよばれる<ref name="Beakes2014" /><ref name="Beakes2017" />。 |
|||
卵菌の初期分岐群の特徴から、卵菌の原始形質は海産で単純な[[全実性]]、内部寄生性であると考えられており、このような状態から、core oomycetes の共通祖先において淡水・陸上への進出、発達した菌糸体や腐生性の獲得が起こったとも考えられている<ref name="Beakes2017" />。ただし、core oomycetes の中にも海産、全実性、内部寄生性の種は存在し、このような方向への進化および逆方向への進化が何度も独立に起こったことが示唆されている<ref name="Beakes2017" />。また、有性生殖における配偶子嚢・配偶子の雌雄分化も core oomycetes に限られているが、同型配偶子嚢接合の種も知られており、このような雌雄分化はミズカビ群とツユカビ群で独立に起こった可能性も示唆されている<ref name="Beakes2017" />。ツユカビ群の中では、陸上植物絶対寄生性への進化が、シロサビキン目とツユカビ目において独立に起こったと考えられている<ref name="Beakes2017" />。 |
|||
ツユカビ目は、植物絶対寄生性のツユカビ目(狭義)と腐生能をもつフハイカビ目に分けることもある<ref name="辞典2013分類表" /><ref name="Spring2012">{{cite book|author=Spring, O.|year=2012|chapter=8. Oomycota|editor=Frey, W. (eds.)|title=Syllabus of Plant Families. A. Engler's Syllabus der Pflanzenfamilien Part 1/1|publisher=Borntraeger|isbn=978-3-443-01061-4|pages=98-105}}</ref>。しかし分子系統学的研究によって両群は連続的て明瞭に分けられないことが示されており、2023年現在ではふつうツユカビ目(広義)にまとめられている<ref name="Beakes2017" />。また、ツユカビ目の中でツユカビ科とフハイカビ科に分けられているが、分子系統学的研究に基づいて、後者から前者へ移された例([[エキビョウキン|エキビョウキン属]] {{Snamei||Phytophthora}} など)も多い<ref name="Beakes2017" />。 |
|||
{{cladogram |
|||
|caption='''10'''. 卵菌綱の系統仮説の一例<ref name="Beakes2017" /><ref name="Buaya2021a" /><ref name="Buaya2021b" /><ref name="Buaya2020">{{Cite journal|author=Buaya, A. T. & Thines, M.|year=2020|title=An overview on the biology and phylogeny of the early-diverging oomycetes|journal=Philippine Journal of Systematic Biology |volume=14|issue=|pages=1-20|doi=10.26757/pjsb2020a14004}}</ref><ref name="Marco2016">{{Cite journal|author=Marco, T. & Young-Joon, C.|year=2016|title=Evolution, Diversity, and Taxonomy of the Peronosporaceae, with Focus on the Genus ''Peronospora''|journal=Phytopathology|volume=106|issue=1|pages=6-18|doi=10.1094/PHYTO-05-15-0127-RVW}}</ref><ref name="Tsai2023">{{Cite journal|author=Tsai, I. & Thines, M.|year=2023|title=Adding a missing piece to the puzzle of oomycete phylogeny: the placement of ''Rhipidium interruptum'' (Rhipidiaceae)|journal=Fungal Systematics and Evolution|volume=11|issue=1|pages=95-108|doi=10.3114/fuse.2023.11.08}}</ref> |
|||
|align=center |
|||
|width= |
|||
|clades={{clade| style=font-size:80%;line-height:100% |
|||
|grouplabel1=Basal oomycetes |
|||
|label1='''卵菌綱''' |
|||
|1={{clade |
|||
|1={{clade |
|||
|1=ユーリカスマ目 {{Sname||Eurychasmales}}|barbegin1=green |
|||
|2=ハプトグロッサ目 {{Sname||Haptoglossales}}|bar2=green |
|||
}} |
|||
|2=フクロカビモドキ目 {{Sname||Olpidiopsidales}}|bar2=green |
|||
|3={{Sname||Miraculales}}|bar3=green |
|||
|4=サカゲフクロカビ目 {{Sname||Anisolpidiales}}|bar4=green |
|||
|5={{Sname||Pontismatales}}|bar5=green |
|||
|6=ハリフソロス目 {{Sname||Haliphthorales}}|barend6=green |
|||
|sublabel7= Core oomycetes |
|||
|7={{clade |
|||
|label1=ミズカビ亜綱 |
|||
|1={{clade |
|||
|1=フシミズカビ目 {{Sname||Leptomitales}} |
|||
|2=ミズカビ目 {{Sname||Saprolegniales}} |
|||
}} |
|||
|label2=ツユカビ亜綱 |
|||
|2={{clade |
|||
|1=オオギミズカビ目 {{Sname||Rhipidiales}} |
|||
|2={{clade |
|||
|1=シロサビキン目 {{Sname||Albuginales}} |
|||
|2=ツユカビ目 {{Sname||Peronosporales}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
}} |
|||
<span id="system"></span> |
|||
{| class="wikitable" style="margin:0 auto" |
|||
|'''表3'''. 卵菌の属までの分類体系の一例<ref name="徳増2005">{{cite book|author=徳増征二|year=2005|chapter=卵菌門|editor=杉山純多|title=バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統|publisher=裳華房|isbn=978-4785358273|pages=191–197}}</ref><ref name="バイオ2005">{{cite book|author=|year=2005|chapter=分類表 卵菌門|editor=杉山純多|title=バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統|publisher=裳華房|isbn=978-4785358273|pages=397–398}}</ref><ref name="井上1983" /><ref name="辞典2013分類表">{{cite book|author=|year=2013|chapter=生物分類表|editor=巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一|title=岩波 生物学辞典 第5版|publisher=岩波書店|isbn=978-4000803144|pages=1653–1654}}</ref><ref name="MycoBank">{{Cite web|url=https://www.mycobank.org/|title=The MycoBank|publisher=Robert, V., Stegehuis, G. & Stalpers, J. |accessdate=2023-08-12}}</ref><ref name="Vilela2019">{{Cite journal|author=Vilela, R., Humber, R. A., Taylor, J. W. & Mendoza, L.|year=2019|title=Phylogenetic and physiological traits of oomycetes originally identified as ''Lagenidium giganteum'' from fly and mosquito larvae|journal=Mycologia|volume=111|issue=3|pages=408-422|doi=10.1080/00275514.2019.1589316}}</ref><ref name="Duffy2015">{{Cite journal|author=Duffy, M. A., James, T. Y. & Longworth, A.|year=2015|title=Ecology, virulence, and phylogeny of ''Blastulidium paedophthorum'', a widespread brood parasite of ''Daphnia'' spp.|journal=Applied and Environmental Microbiology|volume=81|issue=16|pages=5486-5496|doi=10.1128/AEM.01369-15}}</ref><ref name="Buaya2020" /><ref name="Buaya2021a">{{Cite journal|author=Buaya, A.T., Scholz, B. & Thines, M.|year=2021|title=Taxonomy and phylogeny of ''Aphanomycopsis bacillariacearum'', a holocarpic oomycete parasitoid of the freshwater diatom genus ''Pinnularia''|journal=Mycol Progress|volume=20|issue=|pages=289–298|doi=10.1007/s11557-021-01668-x}}</ref><ref name="Tsai2023" /> |
|||
* 偽菌門 {{Sname||Pseudofungi}} {{AUY|Caval.-Sm.|1989|bio=bot}}<br /><small>または</small>卵菌門 {{Sname||Oomycota}} {{AUY|Arx|1967|bio=bot}} |
|||
** 卵菌綱 {{Sname||Oomycetes}} {{AUY|G. Winter|1880|bio=bot}} [syn. ツユカビ綱 (広義) {{Sname||Peronosporomycetes}} {{AUY|Locq.|1974|bio=bot}} ''sensu lato''](複数の綱に分けることもある; 上記参照) |
|||
*** ユーリカスマ目 {{Sname||Eurychasmales}} {{AU|Sparrow}} ex {{AUY|A. Buaya & Thines|2020|bio=bot}}<ref name="Buaya2020" /> |
|||
**** ユーリカスマ科 {{Sname||Eurychasmataceae}} {{AUY|H.E. Petersen|1905|bio=bot}} |
|||
****:[[全実性]]、[[褐藻]]に寄生。 |
|||
***** {{Snamei||Eurychasma}}, {{Snamei||Eurychasmidium}} |
|||
*** ハプトグロッサ目 {{Sname||Haptoglossales}} {{AUY|M.W. Dick|2001|bio=bot}} |
|||
**** ハプトグロッサ科 {{Sname||Haptoglossaceae}} {{AUY|M.W. Dick|2001|bio=bot}} |
|||
****:全実性、[[線虫]]や[[ワムシ]](ほとんどは土壌生、一部は海産)に寄生、特異な"銃細胞"によって宿主に侵入する。 |
|||
***** {{Snamei||Haptoglossa}} |
|||
*** フクロカビモドキ目 {{Sname||Olpidiopsidales}} {{AUY|M.W. Dick|2001|bio=bot}} |
|||
**** フクロカビモドキ科 {{Sname||Olpidiopsidacae}} {{AU|Sparrow}} ex {{AUY|Cejp|1959|bio=bot}} |
|||
****:全実性、卵菌に寄生する。 |
|||
***** フクロカビモドキ属 {{Snamei||Olpidiopsis}} |
|||
*** {{Sname||Miraculales}} {{AUY|Buaya & Thines|2021|bio=bot}} |
|||
**** {{Sname||Miraculaceae}} {{AUY|Buaya, Hanic & Thines|2017|bio=bot}} |
|||
****:全実性、[[珪藻]]に寄生。 |
|||
***** {{Snamei||Miracula}} |
|||
*** {{Sname||Pontismatales}} {{AUY|Thines|2019|bio=bot}} |
|||
**** {{Sname||Pontismataceae}} {{AU|H.E. Petersen}} ex {{AUY|P.M. Kirk, P.F. Cannon & J.C. David|2001|bio=bot}} |
|||
****:全実性、[[紅藻]]・[[緑藻]]に寄生。 |
|||
***** {{Snamei||Petersenia}}, {{Snamei||Pontisma}} [syn. シロルピジウム属 {{Snamei|Sirolpidium}}]<ref name="Buaya2021b">{{Cite journal|author=Buaya, A. T., Scholz, B. & Thines, M.|year=2021|title=''Sirolpidium bryopsidis'', a parasite of green algae, is probably conspecific with ''Pontisma lagenidioides'', a parasite of red algae|journal=Fungal Systematics and Evolution|volume=7|issue=1|pages=223-231.|doi=10.3114/fuse.2021.07.11}}</ref> |
|||
*** サカゲフクロカビ目 {{Sname||Anisolpidiales}} {{AUY|M.W. Dick|2001|bio=bot}} |
|||
**** サカゲフクロカビ科 {{Sname||Anisolpidiaceae}} {{AU|Karling}} ex {{AUY|M.W. Dick|2001|bio=bot}} |
|||
****:全実性、褐藻に寄生。 |
|||
***** サカゲフクロカビ属 {{Snamei||Anisolpidium}} |
|||
**** {{Sname||Diatomophthoraceae}} {{AUY|Buaya & Thines|2019|bio=bot}} |
|||
****:全実性、[[珪藻]]に寄生。 |
|||
***** {{Snamei||Diatomophthora}} |
|||
*** ハリフソロス目 {{Sname||Haliphthorales}} {{AUY|Beakes, D. Honda & Thines|2014|bio=bot}} |
|||
**** ハリフソロス科 {{Sname||Haliphthoraceae}} {{AUY|Vishniac|1958|bio=bot}} |
|||
****: 分岐・分節化する全実性、[[甲殻類]]などに寄生。 |
|||
***** {{Snamei||Haliphthoros}}, {{Snamei||Halioticida}}, {{Snamei||Halocrusticida}}, {{Snamei||Halodaphnea}} |
|||
*** ミズカビ亜綱 {{Sname||Saprolegniomycetidae}} {{AUY|M.W. Dick|2001|bio=bot}} [ミズカビ綱 {{Sname||Saprolegniomycetes}} {{AUY|R.T. Moore|1994|bio=bot}}] |
|||
***:ときに[[遊走子]]の二回遊泳性を示し、遊走子は放出時に包嚢には包まれない。遊走子および卵形成は遠心的な分割による。生卵器はときに多卵性、卵形成時に周辺細胞質は形成されない。[[ステロール]]合成能をもつ。アンモニウムや有機硫黄を好む。 |
|||
****フシミズカビ目 {{Sname||Leptomitales}} {{AUY|Kanouse|1927|bio=bot}} [syn. アトキンシエラ目 {{Sname||Atkinsiellales}} {{AUY|Beakes, D. Honda & Thines|2014|bio=bot}}]<ref name="Buaya2020c" /> |
|||
***** アトキンシエラ科 {{Sname||Atkinisellaceae}} {{AUY|Sparrow|1976|bio=bot}} |
|||
*****:菌糸状で[[全実性]]、[[甲殻類]]に寄生。 |
|||
****** {{Snamei||Atkinsiella}} |
|||
***** クリプチコラ科 {{Sname||Crypticolaceae}} {{AUY|M.W. Dick|1995|bio=bot}} |
|||
*****:菌糸状で全実性、[[双翅目]]幼虫に寄生。 |
|||
****** {{Snamei||Crypticola}} |
|||
***** ラゲニスマ科 {{Sname||Lagenismataceae}} {{AUY|M.W. Dick|1995|bio=bot}} |
|||
*****:菌糸状で全実性、[[珪藻]]に寄生。 |
|||
****** {{Snamei||Lagenisma}} |
|||
***** エクトゲラ科 {{Sname||Ectrogellaceae}} {{AUY|Cejp|1959|bio=bot}} |
|||
*****:全実性、珪藻に寄生。 |
|||
****** {{Snamei||Ectrogella}} [syn. {{Snamei|Cymbanche}}] |
|||
***** レプトレグニエラ科 {{Sname||Leptolegniellaceae}} {{AUY|M.W. Dick|1971|bio=bot}} [syn. {{Sname|Ducellieriaceae}} {{AUY|M.W. Dick|2001}}] |
|||
*****:全実性から菌糸体性(菌糸はしばしばくびれる)、腐生性または藻類などに寄生。 |
|||
****** {{Snamei||Aphanomycopsis}}, {{Snamei||Brevilegniella}}, {{Snamei||Cornumyces}}, {{Snamei||Ducellieria}}, {{Snamei||Eurychasmopsis}}, {{Snamei||Leptolegniella}}, {{Snamei||Nematophthora}}, {{Snamei||Pythiella}} |
|||
***** フシミズカビ科 {{Sname||Leptomitaceae}} {{AUY|Kütz.|1843|bio=bot}} [syn. アポダクリエラ科 {{Sname|Apodachlyellaceae}} {{AUY|M.W. Dick|1986}}] |
|||
*****:全実性または菌糸体性(菌糸はしばしばくびれる)、腐生性(ときに嫌気的環境)または甲殻類に寄生。 |
|||
****** {{Snamei||Apodachlya}}, {{Snamei||Apodachlyella}}, {{Snamei||Blastulidium}}, フシミズカビ属 {{Snamei||Leptomitus}}, {{Snamei||Periplasma}} |
|||
***** 所属不明: {{Snamei||Chlamydomyzium}}([[線虫]]に寄生), {{Snamei||Bolbea}}([[カイミジンコ]]に寄生) |
|||
**** ミズカビ目 {{Sname||Saprolegniales}} {{AUY|E. Fisch.|1892|bio=bot}} |
|||
***** ベルカルブス科 {{Sname||Verrucalvaceae}} {{AUY|M.W. Dick|1984|bio=bot}} |
|||
*****:菌糸体性で菌糸や遊走子嚢が細い、腐生性または植物の根や線虫、甲殻類などに寄生。 |
|||
****** {{Snamei||Aphanomyces}}, {{Snamei||Pachymetra}}, {{Snamei||Plectospira}}, {{Snamei||Sommerstorffia}}, {{Snamei||Verrucalvus}} |
|||
***** [[ミズカビ|ミズカビ科]] {{Sname||Saprolegniaceae}} {{AU|Kütz.}} ex {{AUY|Warm.|1884|bio=bot}}{{efn2|name="ミズカビ科"|ワタカビ属、アミワタカビ属、ヤブレワタカビ属、{{Snamei||Brevilegnia}} をワタカビ科 ("{{Sname|Achlyaceae}}") として分けることも提唱されている<ref name="Beakes2014" />。}} [syn. レプトレグニア科 {{Sname|Leptolegniaceae}} {{AUY|M.W. Dick|1999}}] |
|||
*****:菌糸体性で菌糸は太い、ほとんどは腐生性。 |
|||
****** ワタカビ属 {{Snamei||Achlya}}, {{Snamei||Aplanes}}, {{Snamei||Aplanopsis}}, {{Snamei||Brevilegnia}}, {{Snamei||Calyptralegnia}}, {{Snamei||Couchia}}, アミワタカビ属 {{Snamei||Dictyuchus}}, {{Snamei||Geolegnia}}, {{Snamei||Hydatinophagus}}, {{Snamei||Isoachlya}}, {{Snamei||Leptolegnia}}, {{Snamei||Newbya}}, {{Snamei||Protoachlya}}, {{Snamei||Pythiopsis}}, [[ミズカビ|ミズカビ属]] {{Snamei||Saprolegnia}}, {{Snamei||Scoliolegnia}}, ヤブレワタカビ属 {{Snamei||Thraustotheca}} |
|||
***** 所属不明: {{Snamei||Aquastella}}(ワムシに寄生) |
|||
*** ツユカビ亜綱 {{Sname||Peronosporomycetidae}} {{AUY|M.W. Dick|2001|bio=bot}} [ツユカビ綱 (狭義) {{Sname||Peronosporomycetes}} {{AUY|Locq.|1974|bio=bot}} ''sensu stricto''] |
|||
***:遊走子は一回遊泳性であり、しばしば包嚢に包まれて放出される。生卵器はふつう1卵性、球心的に形成され、周辺細胞質を形成する。ステロール合成能を欠く。さまざまな窒素・硫黄源を利用可能(絶対寄生性種を除く)。 |
|||
**** オオギミズカビ目 {{Sname||Rhipidiales}} {{AUY|M.W. Dick|2001|bio=bot}} |
|||
***** {{Sname||Salispinaceae}} {{AUY|R. Benn. & Thines|2018|bio=bot}} |
|||
*****:菌糸体性、汽水、腐生性。 |
|||
****** {{Snamei||Salispina}} |
|||
***** オオギミズカビ科 {{Sname||Rhipidiaceae}} {{AU|Sparrow}} ex {{AUY|Cejp|1959|bio=bot}} |
|||
*****:菌糸体性または基底細胞と仮根からなる特異な菌体、腐生性。 |
|||
****** {{Snamei||Araiosporab}}, {{Snamei||Aqualinderella}}, {{Snamei||Mindeniella}}, {{Snamei||Nellymyces}}, オオギミズカビ属 {{Snamei||Rhipidium}}, {{Snamei||Sapromyces}} |
|||
**** シロサビキン目 {{Sname||Albuginales}} {{AUY|Thines|2005|bio=bot}} |
|||
***** シロサビキン科 {{Sname||Albuginaceae}} {{AUY|J. Schröt.|1857|bio=bot}} |
|||
*****:菌糸体性、維管束植物に寄生。 |
|||
****** シロサビキン属 {{Snamei||Albugo}} [syn. {{Snamei|Cystopus}}], {{Snamei||Pustula}}, {{Snamei||Wilsonia}} |
|||
**** ツユカビ目 {{Sname||Peronosporales}} {{AUY|Bek.|1863|bio=bot}}(広義)[syn. フハイカビ目 {{Sname|Pythiales}} {{AUY|M.W. Dick|2001}}; ササラビョウキン目 {{Sname|Sclerosporales}} {{AUY|M.W. Dick|1986}}; クサリフクロカビ目 {{Sname|Lagenidiales}} {{AUY|Karling|1939}}] |
|||
***** {{Sname||Salisapiliaceae}} {{AUY|Hulvey, Nigrelli, Telle, Lamour & Thines|2010|bio=bot}} |
|||
*****:菌糸体性、汽水、腐生性。 |
|||
****** {{Snamei||Salisapilia}} |
|||
***** フハイカビ科 {{Sname||Pythiaceae}} {{AUY|J. Schröt.|1897|bio=bot}} [syn. ピチオゲトン科 {{Sname|Pythiogetonaceae}} {{AUY|M.W. Dick|1999}}; ラゲナ科 {{Sname|Lagenaceae}} {{AUY|M.W. Dick|2001}}; クサリフクロカビ科 {{Sname|Lagenidiaceae}} {{AUY|J. Schröt.|1897}}; {{Sname|Peronophythoraceae}} {{AUY|W.H. Ko, H.S. Chang, H.J. Su, C.C. Chen & L.S. Leu|1978}}; {{Sname|Myzocytiopsidaceae}} {{AUY|M.W. Dick|1997}} |
|||
*****:全実性から菌糸体性、腐生性または維管束植物、藻類、動物などに寄生。 |
|||
****** {{Snamei||Cystosiphon}} [syn. {{Snamei|Lagenidiopsis}}], {{Snamei||Gominocheate}}, クサリフクロカビ属 {{Snamei||Lagenidium}}, {{Snamei||Lagena}} ({{Snamei|Lagenocystis}}), {{Snamei||Myzocytiopsis}}, {{Snamei||Myzocytium}}, {{Snamei||Salilagenidium}} |
|||
****** {{Snamei||Diasporangium}}, {{Snamei||Elongisporangium}}, {{Snamei||Globisporangium}}, {{Snamei|Lucidium}}, {{Snamei||Peronophythora}}, {{Snamei||Pilasporangium}}, {{Snamei||Pythiogeton}} [syn. {{Snamei|Plasmoparopsis}}], [[フハイカビ|フハイカビ属]](ピシウム) {{Snamei||Pythium}} [syn. {{Snamei|Artotrogus}}, {{Snamei|Eupythium}}, {{Snamei|Nematosporangium}}, {{Snamei|Rheosporangium}}], {{Snamei||Trachysphaera}} |
|||
***** ツユカビ科 {{Sname||Peronosporaceae}} {{AUY|de Bary|1863|bio=bot}} [syn. エキビョウキン科 {{Snamei|Phytophthoraceae}} {{AUY|Haeckel|1894|bio=bot}}; ササラビョウキン科 {{Sname|Sclerosporaceae}} {{AUY|M.W. Dick|1984}}] |
|||
*****:菌糸体性、腐生性または維管束植物に寄生。 |
|||
****** {{Snamei||Baobabopsis}}, スリコギツユカビ属 {{Snamei||Basidiophora}} ({{Snamei|Gilletia}}), {{Snamei||Benua}}, ラッパツユカビ {{Snamei||Bremia}} ({{Snamei|Actinobotrys}}, {{Snamei|Tetradium}}), {{Snamei||Calycofera}}, {{Snamei||Eraphthora}}, {{Snamei||Graminivora}}, {{Snamei||Halophytophthora}}, {{Snamei||Hyaloperonospora}}, {{Snamei||Nothophytophthora}}, {{Snamei||Novotelnova}}, {{Snamei||Paraperonospora}}, {{Snamei||Perofascia}}, {{Snamei||Peronosclerospora}}, ツユカビ属 {{Snamei||Peronospora}} ({{Snamei|Bremiella}}), [[エキビョウキン|エキビョウキン属]] {{Snamei||Phytophthora}} [syn. {{Snamei|Blepharospora}}, {{Snamei|Kawakamia}}, {{Snamei|Mycelophagus}}, {{Snamei|Nozemia}}, {{Snamei|Phloeophthora}}, {{Snamei|Pythiacystis}}, {{Snamei|Pythiomorpha}}, {{Snamei|Sphaerosporangium}}], {{Snamei||Phytopythium}} [syn. {{Snamei|Ovatsporangium}}], タンジクツユカビ属 {{Snamei||Plasmopara}} ({{Snamei|Pseudoplasmopara}}, {{Snamei|Rhysotheca}}), {{Snamei||Plasmoverna}}, {{Snamei||Poakatesthia}}, {{Snamei||Protobremia}}, ニセツユカビ属 {{Snamei||Pseudoperonospora}} ({{Snamei|Peronoplasmopara}}), {{Snamei||Scleropthora}}, ササラビョウキン属 {{Snamei||Sclerospora}}, {{Snamei||Synchrospora}}, {{Snamei||Viennotia}} |
|||
***** 所属不明: {{Snamei||Paralagenidium}}(哺乳類に寄生) |
|||
*** 所属不明 |
|||
**** ロゼロプシス科 {{Sname||Rozellopsidaceae}} {{AUY|M.W. Dick|1995|bio=bot}}: {{Snamei||Rozellopsis}} [syn. {{Snamei|Skirgiellopsis}}] |
|||
**** プセウドスファエリタ科 {{Sname||Pseudosphaeritaceae}} {{AUY|M.W. Dick|1995|bio=bot}}: {{Snamei||Pseudosphaerita}}, {{Snamei||Sphaerita}} |
|||
**** {{Snamei||Calycofera}}, {{Snamei||Ciliatomyces}} [syn. {{Snamei|Ciliomyces}}], {{Snamei||Olpidiomorpha}}, {{Snamei||Pleotrachelus}}, {{Snamei||Bicilium}}, {{Snamei||Diplophysa}}, {{Snamei||Peronium}}, {{Snamei||Pleocystidium}}, {{Snamei||Pseudolpidiopsis}}, {{Snamei||Pseudolpidium}} |
|||
|} |
|||
=== 化石記録 === |
|||
卵菌類と考えられる化石は、古生代[[デボン紀]]前期の[[ライニーチャート]]から報告されている<ref name="Beakes2017" />。また、[[石炭紀]]の地層からは、[[リンボク (化石植物)|リンボク]]([[小葉植物]])や[[シダ植物]]、[[裸子植物]]に寄生していた可能性がある卵菌の卵胞子や生卵器の化石が報告されている<ref name="Beakes2017" /><ref>{{Cite journal|author=Dotzler, N., Krings, M., Agerer, R., Galtier, J. & Taylor, T. N.|year=2008|title=''Combresomyces cornifer'' gen. sp. nov., an endophytic peronosporomycete in ''Lepidodendron'' from the Carboniferous of central France|journal=Mycological Research|volume=112|issue=9|pages=1107-1114|doi=10.1016/j.mycres.2008.03.003}}</ref><ref>{{Cite journal|author=Harper, C. J., Krings, M., Galtier, J. & Taylor, T. N.|year=2016|title=A microfossil with suggested affinities to the Peronosporomycetes Oomycota from the Carboniferous c. 330 Ma of France|journal=Nova Hedwig|volume=103|issue=3-4|pages=315-326|doi=10.1127/nova_hedwigia/2016/0352}}</ref><ref>{{Cite journal|author=Strullu-Derrien, C., Gèze, M., Spencer, A. R., De Franceschi, D., Kenrick, P., Selosse, M. A. & Knoll, A. H.|year=2021|title=An expanded diversity of oomycetes in Carboniferous forests: Reinterpretation of Oochytrium lepidodendri (Renault 1894) from the Esnost chert, Massif Central, France|journal=Plos One|volume=16|issue=3|pages=e0247849|doi=}}</ref>。また、[[白亜紀]]の[[琥珀]]からも卵菌類と考えられる構造が見つかっている<ref>{{cite web|url=http://www.ucmp.berkeley.edu/chromista/oomycota.html|title=Introduction to the Oomycota|accessdate=2012-09-}}</ref>。 |
|||
== ギャラリー == |
|||
<gallery style="font-size:80%;"> |
|||
File:White rust symptoms.jpg|{{Snamei||Albugo tragopogonis}}(シロサビキン目)が寄生した[[ヒマワリ]] |
|||
File:Pythium 5361373.png|[[フハイカビ|フハイカビ属]](ツユカビ目)が寄生した[[インゲンマメ]]の根 |
|||
File:Phytophthora blight of papaya (5794539067).jpg|{{Snamei||Phytophthora blight}}(ツユカビ目)が寄生した[[パパイヤ]] |
|||
File:Phytophthora infestans potato Parel, aardappelziekte Parel.jpg|ジャガイモエキビョウキン(ツユカビ目)が寄生した[[ジャガイモ]] |
|||
File:Peronospora manshurica on soybean leaf.jpg|{{Snamei||Peronospora manshurica}}(ツユカビ目)が寄生したダイズ |
|||
File:Draba verna kz20.jpg|{{Snamei||Hyaloperonospora}}(ツユカビ目)が寄生したヒメナズナ |
|||
File:Hyaloperonospora-parasitica-hyphae-haustoria.jpg|[[シロイヌナズナ]]に寄生した {{Snamei||Hyaloperonospora}}(ツユカビ目)の菌糸と植物細胞内に侵入させた吸器(トリパンブルー染色試料) |
|||
File:Nsr-slika-329.png|ツユカビ属(ツユカビ目)の菌糸体と気孔から伸ばした胞子嚢柄 |
|||
File:Pseudoperonospora cubensis sporangiophore.jpg|{{Snamei||Pseudoperonospora cubensis}}(ツユカビ目)の胞子嚢柄 |
|||
File:Pseudoperonospora farinosa f.sp. spinaciae at Spinacia oleracea sporangiën (3).jpg|{{Snamei||Pseudoperonospora}}(ツユカビ目)の遊走子嚢 |
|||
File:10554 oospore.jpg|[[エキビョウキン|エキビョウキン属]](ツユカビ目)の卵胞子 |
|||
File:Oospore of Phytophthora infestans.jpg|エキビョウキン属(ツユカビ目)の卵胞子 |
|||
</gallery> |
|||
== 脚注 == |
== 脚注 == |
||
{{脚注ヘルプ}} |
|||
=== 注釈 === |
|||
{{Notelist2}} |
|||
=== 出典 === |
|||
{{Reflist}} |
{{Reflist}} |
||
== 外部リンク == |
|||
{{Commonscat|Oomycetes}} |
|||
{{wikispecies|Oomycetes}} |
|||
* {{Cite web|url=https://www.mycobank.org/|title=The MycoBank|publisher=Robert, V., Stegehuis, G. & Stalpers, J. |accessdate=2023-09-22}} (英語) |
|||
* {{Cite web|author=Fry, William E. & Grünwald, Niklaus J.|date=2010|url=https://www.apsnet.org/edcenter/disandpath/oomycete/introduction/Pages/IntroOomycetes.aspx|title=Introduction to Oomycetes|website=|publisher=The American Phytopathological Society|doi=10.1094/PHI-I-2010-1207-01|accessdate=2023-09-23}} (英語) |
|||
{{デフォルトソート:らんきん}} |
{{デフォルトソート:らんきん}} |
||
[[Category:ストラメノパイル]] |
[[Category:ストラメノパイル]] |
||
2023年10月7日 (土) 21:21時点における版
| 卵菌 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 1. ワタカビ属(ミズカビ目)の菌糸と卵胞子
| |||||||||||||||||||||
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 学名 | |||||||||||||||||||||
| Oomycota Arx (1967) | |||||||||||||||||||||
| シノニム | |||||||||||||||||||||
| |||||||||||||||||||||
| 英名 | |||||||||||||||||||||
| oomycete | |||||||||||||||||||||
| 下位分類 | |||||||||||||||||||||
卵菌(らんきん、英: oomycetes)とは、ストラメノパイルに属する吸収栄養性(細胞膜を通して低分子有機物を吸収する)の原生生物の一群、またはこれに属する生物のことであり、その形態や生活様式は菌類に似ているが、系統的には全く異なる。微小な単細胞性のものから、発達した菌糸を形成するものまであり、菌糸は基本的に隔壁を欠く(図1)。セルロースを含む細胞壁をもつ。多くは、2本の不等鞭毛をもつ遊走子による無性生殖を行い、配偶子嚢接合による有性生殖を行う(図1)。栄養体は複相(染色体を2セットもつ)であり、配偶子嚢内で減数分裂を行う。海や淡水、陸上に広く分布しており、腐生性(生物遺体など生きていない有機物を分解して栄養を吸収する)、または陸上植物や藻類、動物などに寄生する。植物病原菌となるものが多く知られており、ジャガイモエキビョウキン(ツユカビ目)は19世紀にアイルランドでジャガイモ飢饉を引き起こし、多数の餓死者と北米への移民の原因となった。
100属1,700種ほどが知られる大きなグループであり、分類学的には、卵菌綱(学名: Oomycetes)にまとめられることが多いが、独自の卵菌門(学名: Oomycota)にまとめて複数の綱に分けることもある。菌類に似ているため、古くは菌類の中の藻菌類や鞭毛菌類に分類されていた。しかし卵菌と菌類は系統的に非常に離れており、この類似性は収斂進化によるものである。
特徴
体制
菌体の体制はさまざまであり、単細胞で全実性(全体が遊走子嚢など生殖器官になる)のものから、発達した菌糸を形成するもの(下図2)まである[1][2][3][4]。全実性のものでは、単純な球状の種から、菌糸状に細長いものや、分岐・分節化した種まである[1][3]。卵菌の中で初期分岐群は全実性・寄生性であるが、ミズカビ目やツユカビ目など派生的な群では発達した菌糸をもつものが多い[3][5][6]。ただし派生的な群の中にも、全実性・寄生性である種が含まれる。菌糸の太さはさまざまであり、直径2マイクロメートル (µm) 程度のものから、150 µm に達するものまである[3]。菌糸は先端成長を行い、ときに分枝し、ふつう隔壁を欠く無隔菌糸であるが(下図2)、古くなった部分や生殖器を区切る隔壁が形成されることもある[1][3]。菌糸性で絶対寄生性の種の多くは、宿主細胞間に菌糸を伸ばし、宿主細胞内に吸器を挿入する[1][3](下図2c)。



菌体はふつう細胞壁に包まれるが、寄生性の初期分岐群の中には、栄養体(通常時の体)が宿主内で細胞壁を欠く多核細胞(変形体)として生きている例もある[2][3]。卵菌の細胞壁はおもにセルロースとβ-グルカンからなる。ただし一部の種からはキチンも報告されており、またゲノムの調査からは全ての卵菌類がキチン合成酵素をもつことが示唆されている[7]。フシミズカビ目では、菌糸のくびれ部分にキチンを含むセルリン顆粒が存在することがある[3]。
細胞
卵菌の栄養体は、基本的に多数の核を含む多核性 (coenocytic) である[1][2][3]。核分裂は核膜が維持される閉鎖型であり、核内に紡錘体が形成され、両極には中心体が存在する[1][3]。
栄養体中央はふつう液胞で占められており、核や他の細胞小器官を含む細胞質は周縁部に位置する[3]。ミトコンドリアのクリステは管状である[1][3]。多くの菌類とは異なり、典型的なゴルジ体をもつ[1][3]。貯蔵多糖はリン酸化した水溶性のβ-1,3グルカンであり、マイコラミナリン (mycolaminarin) とよばれる[1]。マイコラミナリンは小胞に包まれてときに層状構造を示し、この小胞は dense-body vesicle または fingerprint vacuole ともよばれる[1]。菌類に一般的な貯蔵多糖であるグリコーゲン顆粒は、卵菌には見られない[1]。また、ポリリン酸体を欠く[1]。アミノ酸のリシン合成経路はジアミノピメリン酸経路(DAP経路)であり、菌類に見られるα-アミノアジピン酸経路(AAA経路)とは異なる[1]。
無性生殖
卵菌は、基本的に遊走子(鞭毛をもつ胞子)による無性生殖を行う(下図3a)。菌体の全体または一部が遊走子嚢となり、細胞壁の一部が溶解することで遊走子を放出する[3][4]。ただし鞭毛を欠く不動胞子 (aplanospore) を放出することもあり、また鞭毛形成能を完全に欠いている種もいる[3]。遊走子は好適な環境に着生すると、再び菌体へと成長する[1][3]。全実性で寄生性の種では、放出される遊走子の通り道となる逸出管を形成することが多い[3]。菌糸を形成するものでは、ふつう菌糸先端が隔壁によって区切られて遊走子嚢を形成する[3](下図3b左)。シロサビキン属(シロサビキン目)では、遊走子嚢が鎖状に連なって形成される[3](下図3b中)。ツユカビ属(ツユカビ目)などでは、有限成長する分枝した遊走子嚢柄 (sporangiophore) の枝先に遊走子嚢をつける[3](下図3b右, 3c)。
卵菌の初期分岐群やミズカビ亜綱では、遊走子形成は遠心的であり、また細胞分裂が完了したのちに鞭毛形成が起こる[3]。それに対してツユカビ亜綱では、遊走子形成は求心的であり、また遊走子の分化と鞭毛形成は同時に起こる[3]。一部の卵菌では、遊走子嚢から放出された遊走子または原形質が包嚢 (vesicle) で包まれ、後者の場合はこの中で遊走子への分化が起こる[3]。陸上植物に寄生する種では、しばしば遊走子嚢が切り離されて風や雨で散布され、水の存在によって散布先で遊走子を放出するが、遊走子段階を省略して直接発芽することも多い[1][3](下図3b右, 6)。このように直接発芽する遊走子嚢は、分生子 (conidium) ともよばれる[2][3]。
.png)


遊走子は、基本的に細胞腹面(側面)から生じて前後に伸びる2本の不等鞭毛をもつ[1](上図3d(b))。例外的に、サカゲフクロカビ属(Anisolpidium; サカゲフクロカビ目)では、細胞前端付近から前鞭毛のみが生じている[注 1]。ストラメノパイルに属する他の生物と同様、前鞭毛には管状小毛(管状マスチゴネマ)が付随した羽型鞭毛であり、後鞭毛はこれを欠く尾型鞭毛である[1][2](上図3d)。遊走子が着生する際には、遊走子細胞内に存在する細胞壁物質を含んだ小胞がエクソサイトーシスされて細胞壁を形成してシスト化する[1]。
_198.svg)
ミズカビ亜綱の種は、2本の鞭毛が細胞前端付近から生じている遊走子と、細胞腹面から生じている遊走子をもつことがあり、前者を一次遊走子 (primary zoospore, auxiliary zoospore)、後者を二次遊走子 (secondary zoospore, principal zoospore) とよばれる[1][2][3](上図3d, 5)。このような遊走子の二形性は二回遊泳性 (diplanetism) ともよばれる[1][2][4]。ミズカビ属(Saprolegnia; ミズカビ目)では、菌糸先端の遊走子嚢から放出された一次遊走子は着生、鞭毛を吸収して細胞壁を形成しシスト化し、そこから1個の二次遊走子が生じて再び遊泳する[1][3](下図5)。二次遊走子は鞭毛を落としてシスト化し、菌糸を伸ばして発芽するか、または再び二次遊走子が形成される(数回遊泳性 polyplanetism)[1][3]。ミズカビ目の中には、これを基本形としてさまざまな様式があり、Aphanomyces やワタカビ属 (Achlya) では遊走子嚢の開口部で、アミワタカビ属 (Dictyuchus) やヤブレワタカビ属 (Thraustotheca) では遊走子嚢中で、一次遊走子に相当する細胞がシスト化し、二次遊走子を放出、または直接発芽する[1][3](図4)。一方、Phythiopsis は一次遊走子のみを形成し、これがシスト化して発芽する[1]。
遊走子は、明瞭な細胞外皮をもたない[2]。他のストラメノパイルと同様、遊走子は基本的に4組の微小管性鞭毛根(R1–R4)をもつ[2][3]。鞭毛移行部(鞭毛と基底小体の移行部)には、ふつう2重のらせん構造 (transitional helis) が存在するが、1重らせんのものやこれを欠くものもある[2][3]。遊走子の細胞内にはさまざまなタイプの小胞が存在し、付着や外被形成に関与すると考えられている[2][3](上記参照)。
また、卵菌では菌体の一部が厚壁化し、無性生殖の単位となることがある。この構造は菌芽 (gemma) や厚壁胞子 (chlamydospore) とよばれ、休眠構造として機能し、好適な条件で発芽管を伸ばすか、または遊走子嚢を形成する[1][2][4]。
有性生殖
卵菌の中で生活環における核相変化が知られているものでは、栄養体が複相(染色体を2セットもつ)であり、配偶子形成時に減数分裂を行う[3][4]。ただし初期分岐群では、核相変化・有性生殖が報告されていないものが多い[3]。卵菌の有性生殖では、鞭毛をもつ配偶子の合体は見られず、不動状態の配偶子嚢が直接接着する(配偶子嚢接合 gametangial conjugation)ことで起こる[3]。この結果形成される接合子は卵胞子 (oospore) とよばれ、ふつう休眠細胞となり、発芽して遊走子嚢を形成、または菌糸など栄養体となる[3][4]。
_200.svg)
.png)
Eurychasma(ユーリカスマ目)や Anisolpidium(サカゲフクロカビ目)、Lagenisma(フシミズカビ目)では、鞭毛細胞が着生してシスト化したものが受精管 (fertilization tube) でつながり、一方の原形質がもう一方へ送り込まれ、細胞質・核融合が起きて接合子が形成される[3]。Lagenisma coscinodisciでは、減数分裂によって形成された遊走子が着生して接合することが報告されている[10]。このような雌雄の分化がない同型の配偶子嚢接合が卵菌における原始形質であると考えられている[3]。
ミズカビ亜綱やツユカビ亜綱の多くでは、配偶子嚢の雌雄の分化が見られる[1][3]。雌雄配偶子嚢内では、減数分裂が起こり、単相(染色体を1セットのみもつ)の核を形成する[1][3]。雌性配偶子嚢である生卵器 (oogonium) は、1個から数個の単相の卵(卵球 oosphere ともよばれる)を形成する[1][3][4]。雄性配偶子嚢である造精器 (antheridium) は生卵器に接し(配偶子嚢接着 gametangial contact)、受精管を通じて単相の雄性核を送り込み、卵において受精が起こる[1][3][4]。配偶子嚢や卵胞子の形態は、このような卵菌の分類において極めて重視されている[3]。このような有性生殖は、ステロイド性のホルモンによって制御されていることが知られている[1][3]。ツユカビ亜綱はステロール合成能を欠き、有性生殖のためには外部からステロールを得る必要がある[1][3]。
有性生殖は、ホモタリック(雌雄同株)のものと、ヘテロタリック(雌雄異株)のものが知られている[3]。ホモタリックの場合、生卵器と造精器のできる位置関係には多様性があり、造精器が生卵器の直下にできるもの (直下性 hypogynous)、生卵器と同一の柄に側生するもの (雌雄同菌糸性 monoclinous)、同一の菌体の異なる菌糸にできるもの (雌雄異菌糸性 diclinous) がある[3][11]。エキビョウキン属(ツユカビ目)では、生卵器が造精器を貫通して発達し、生卵器の基部を造精器が取り巻いている状態 (amphigynous) になるものが多い[3](上図6)。また、ヘテロタリックの場合、相手によって雌雄性が切り替わる相対的雌雄性 (relative sexuality) を示すものも知られており、相手が生卵器を形成する場合は造精器を、相手が造精器を形成する場合は生卵器を形成する[1]。また一部の種では、造精器と接着せずに、1個の生卵器の中で2個の単相核が合体することや、減数分裂せずに複相の核をもとに卵胞子が形成されることがある[1]。
生卵器内での卵形成様式には多様性があり、ミズカビ亜綱では遠心的に、ツユカビ亜綱では求心的に卵が形成される[3]。またツユカビ亜綱では、周辺の細胞質が卵にならずに取り残されて周辺細胞質 (periplasm) となり、卵胞子の細胞壁形成などに寄与する[3]。
卵胞子内にはooplastとよばれる大きな液胞(マイコラミナリンやリン酸を含む)が存在し、また周縁部には脂質顆粒(油滴)が存在する[1][3]。このooplastと油滴の配置様式に基づいて下記のようにいくつかの型に分けられており、分類形質として用いられている[1][3][12]。
- セントリック (中心生、centric) … ooplastが中央に位置しており、その周囲を1–2層の小脂質顆粒が囲んでいる。
- サブセントリック (亜中心生、subcentric) … ooplastがやや偏って位置しており、周辺の小脂質顆粒が多層の部分と1–0層の部分がある。
- サブエクセントリック (subeccentric) … ooplastが偏って位置しており、その反対側に複数の脂質顆粒がある。
- エクセントリック (偏心生、eccentric) … ooplastが偏って位置しており、その反対側に1個の大きな脂質顆粒がある。
卵胞子は厚くふつう多層の細胞壁に囲まれており、また突起などで装飾されていることもある[3]。卵胞子の細胞壁内層はendospore、外層はepisporeとよばれる[1]。卵胞子は耐久構造となり、ふつう休眠ののちに発芽して発芽管を伸ばす。ふつう発芽管の先端に遊走子嚢を形成するが、直接菌糸を伸ばすことや、卵胞子内で遊走子が形成されることもある[3]。
生態
卵菌は海や淡水、陸上に広く分布する[1][3]。栄養様式としては、腐生性のものと寄生性のものがいる。
腐生性の種は、植物遺体や動物遺体を分解する[3][4](下図7a, b)。淡水域には、ミズカビ目の腐生性の種が極めてふつうに見られる[3]。一般的に、このような卵菌は湖沼の沿岸域に多く、四季がある地域では春と秋にピークがある[3]。また海の沿岸域でも、Halophytophthora や Salisapilia(ツユカビ目)が、マングローブ林や塩性湿地の植物質の初期分解に重要な役割を担っている[3]。
カップに水サンプルまたは水と土壌サンプルや水中の枯れ枝などを入れ、これに基質となるアサの実やハエの死骸などを加えておくことによって、数日で腐生性の卵菌、特にミズカビ科の菌糸が見られるようになる(このような手法は釣菌法(ちょうきんほう)とよばれる)[1][13]。
卵菌の多くは好気性であるが、フシミズカビ目やオオギミズカビ目、ツユカビ目の中には、嫌気的水域で有機酸発酵などによって生育する通性嫌気性のものも知られている[1][3]。
寄生性の種の中には、宿主の一部を殺して分解・吸収する殺生栄養性のものと、宿主を生かした状態で栄養を吸収する活物栄養性のものがある[1][2][3][4]。腐生性と殺生栄養性の間は連続的であり、条件によって変わるものもいる[1][2]。このような条件的寄生性卵菌の中には、Aphanomyces(ミズカビ目)、フハイカビ属(Pythium)、エキビョウキン属(Phytophthora)(ツユカビ目)など土壌生態系において分解者としても重要な存在もいる[3]。活物栄養性の種は、ふつう絶対寄生性(寄生しなければ生きられない)である[1]。
寄生性の種は、陸上植物に寄生するものが多く知られており、組織の細胞間に菌糸を伸ばしている[3][4](上図2c, 下図7c, d)。宿主はふつう草本性の被子植物であるが、木本に寄生する種もいる[3]。また、宿主特異性が比較的高いものが多いが、様々な植物に寄生する宿主範囲が広いもの(ツユカビ属など)もいる[3]。




陸上植物以外にも、他の卵菌や菌類、海藻(紅藻、緑藻、褐藻)、微細藻(珪藻、接合藻など)に寄生する卵菌も多く知られている[3][14][15][16][17][18][19][20][21][22]。これらの多くは全実性(細胞全体が遊走子嚢になる)であり、宿主細胞内で生育する。
卵菌の中には動物に寄生するものもおり、宿主として甲殻類、昆虫、線虫、ワムシなどが知られている[1][3]。Haptoglossa(ハプトグロッサ目)では、遊走子がシスト化し、銃細胞 (gun cell) とよばれる特殊な細胞となり、線虫に触れると銃弾状の構造を撃ち込み、これを通って原形質が線虫内に侵入する[3]。
人間との関わり
植物寄生性の卵菌の中には農作物など有用植物に寄生して被害を与える種が多く(下図8)、代表的なものを下表1に示した。ジャガイモエキビョウキン(Phytophthora infestans; ツユカビ目)はジャガイモに寄生し、大きな被害を与えることがある(下図8a)。特に19世紀半ば(1845–1847)にジャガイモを主食としていたアイルランドで大飢饉(ジャガイモ飢饉)を引き起こし、大量の餓死者と北米への移民を生み出したため、1841年から1851年の間にアイルランドの人口は162万人も減少した[23]。また、19世紀のヨーロッパでは、北米から侵入したブドウべと病菌(Plasmopara viticola; ツユカビ目)によってブドウ栽培に大きな被害が生じた[24](下図8b)。その対策として、硫酸銅、生石灰、水の混合液であるボルドー液が使われた[1]。1990年代には、米国カリフォルニア州のマテバシイ属やコナラ属の森林において Phytophthora ramorum によるブナ科樹木の急激な枯死 (オーク突然死 Sudden Oak Death) が起こり、大きな被害が生じた[25][26](下図8c)。また、オーストラリアには Phytophthora cinnamomi が侵入し、森林生態系に多大な損害を与えている[26]。

.jpg)
.jpg)
_Pj_DSC_0774.jpg)
| 目 | 科 | 属 | 病例 |
|---|---|---|---|
| ミズカビ目 | ベルカルブス科 | Aphanomyces | ハクサイ根くびれ病、テンサイ黒根病、ホウレンソウ根腐病など |
| シロサビキン目 | シロサビキン科 | シロサビキン属 Albugo | ダイコン白さび病、ハクサイ白さび病、ワサビ白さび病、サツマイモ白さび病など |
| ツユカビ目 | フハイカビ科 | フハイカビ属 Pythium | イネ苗立枯病、トウモロコシ根腐病、ムギ類褐色雪腐病、シバピシウム病、チューリップピシウム葉枯病、ホウレンソウ立枯病、キャベツピシウム腐敗病、スイカ綿腐病、トマト根腐病、サツマイモ白腐病、ニンジンしみ腐病など |
| Globisporangium | イネ苗立枯病、ムギ類褐色雪腐病、ホウレンソウ立枯病、テンサイ苗立枯病、キャベツ苗立枯病、ブロッコリーピシウム腐敗病、レタスピシウム萎凋病など | ||
| ツユカビ科 | Phytopythium | ブロッコリーピシウム腐敗病、イチゴピシウム根腐病、キウイフルーツ根腐病、トマト苗立枯病、バジル根腐病、キクピシウム立枯病など | |
| エキビョウキン属 Phytophthora | ジャガイモ疫病、キュウリ疫病、レタス疫病、イチゴ疫病、ナシ疫病、ネギ白色疫病、アスパラガス疫病、パイナップル心腐病など | ||
| Sclerophthora | イネ黄化萎縮病、コムギ黄化萎縮病、トウモロコシ褐条べと病など | ||
| ササラビョウキン属 Sclerospora | アワしらが病など | ||
| タンジクツユカビ属 Plasmopara | ブドウべと病、ミツバべと病、ヒマワリべと病など | ||
| ラッパツユカビ属 Bremia | シュンギクべと病、レタスべと病など | ||
| Hyaloperonospora | ハクサイべと病、キャベツべと病など | ||
| Peronosclerospora | トウモロコシべと病、サトウキビしらが病など | ||
| ツユカビ属 Peronospora | タマネギべと病、ホウレンソウべと病、ダイズべと病、ソバべと病、タバコべと病、ブロッコリーべと病、バジルべと病など | ||
| ニセツユカビ属 Pseudoperonospora | キュウリべと病、ホップべと病、タイマべと病、エノキべと病など |
養殖海藻であるアマノリ(紅藻)に寄生して被害を与える卵菌がいくつか知られており、Pontisma porphyrae (= Olpidiopsis porphyrae) は壺状菌病を[30][14]、Pythium porphyrae はあかぐされ病を[31]、それぞれ引き起こす。
ミズカビ属やワタカビ属、Aphanomyces(ミズカビ目)などの卵菌は、淡水養殖魚(特にサケ)に寄生して大きな被害を与えることがある[1]。また、Pythium flevoense(ツユカビ目)は、アユ仔魚の内臓真菌症を引き起こすことが報告されている[32]。
ザリガニに寄生する北米産の Aphanomyces astaci(ミズカビ目)は抵抗性をもたない他地域のザリガニに水カビ症(ザリガニペストともよばれる)を引き起こし(下図9a)、スウェーデンでは在来ザリガニ個体群の99%が消失したともされる[33]。そのため、この卵菌は国際自然保護連合(IUCN)が定める「世界の侵略的外来種ワースト100」にも選定されている[33]。この卵菌は日本でも確認されており、在来ニホンザリガニへの影響が懸念されている[33]。


フハイカビ属の Pythium insidiosum(ツユカビ目)は、ヒトを含む哺乳類に対する日和見的な寄生菌となり、ピシウム症 (pythiosis) とよばれる皮膚病を引き起こす[13][34](上図9b)。また、Paralagenidium karlingii(ツユカビ目)も、イヌに寄生して皮膚炎 (paralagenidiosis) を引き起こす[3]。
系統と分類
上位分類
卵菌は吸収栄養性の従属栄養生物であり、またその形態や生活様式は菌類(真菌)とよく似ている。そのため、20世紀後期までは、卵菌は菌類に含められ、菌類の中では、ツボカビ類などとともに藻菌類(子嚢菌・担子菌以外の真菌)や鞭毛菌類(鞭毛細胞をもつ真菌)に分類されることが多かった[4][23][35][36]。しかし、鞭毛細胞の形態や細胞壁組成、リシン合成経路などさまざまな点で、卵菌類は狭義の菌類と異なることが認識されるようになり(下表2)、20世紀末ごろには菌類とは系統的に全く異なる生物群であると考えられるようになった[1][4][36][37]。卵菌の鞭毛細胞や一部の生化学的な特徴は、不等毛藻(褐藻や珪藻)やラビリンチュラ類、一部の原生動物と共通しており、これらが単系統群を構成していると考えられるようになり、この生物群に対してストラメノパイル (stramenopiles; 不等毛類) の名が提唱された[38]。その直後ころから分子系統学的研究が一般的となり、卵菌とこれらの生物との近縁性、ストラメノパイルの存在が支持された[1][36][37]。2023年現在では、卵菌はストラメノパイルの一群として認識されている。卵菌と菌類の間に見られるさまざまな類似点(菌糸など)は、類似した生き方をするため起こった収斂進化の結果であると考えられている[1]。
| ツボカビ類(菌類) | 卵菌類 | |
|---|---|---|
| 細胞壁主要多糖 | キチン | セルロース |
| 転流炭水化物 | ポリオール、トレハロース | グルコース |
| 貯蔵多糖 | グリコーゲン | マイコラミナリン |
| リシン合成 | AAA経路 | DAP経路 |
| ミトコンドリアクリステ | 板状 | 管状 |
| ミトコンドリアのコドン | UGAはトリプトファン | UGAは終止コドン |
| ステロール | エルゴステロール | フコステロール |
| ニコチン酸合成 | トリプトファンから | C3前駆体から |
| 微小管 | ベンゾイミダゾールおよび グリセオフルビン感受性 |
コルヒチン感受性 |
| 鞭毛 | 細胞後端から後方へ尾型鞭毛 | 細胞前端または側面から 前方へ羽型鞭毛、後方へ尾型鞭毛 |
分子系統学的研究から、ストラメノパイルの中で、卵菌はサカゲツボカビ類の姉妹群であることが示唆されている[40][41]。また、珪藻細胞内に侵入する捕食者であるピルソニア類や、自由生活性鞭毛虫であるデヴェロパイエラ類が、卵菌類+サカゲツボカビ類に近縁であることが示唆されている[41]。分類学的には、しばしばこれらを合わせて偽菌門 (Pseudofungi) に分類する[41][42]。ストラメノパイル内において、偽菌門の姉妹群は、不等毛藻(オクロ植物門)であると考えられている[41]。ストラメノパイルの中で菌類的な生物群として、他にラビリンチュラ類があるが(ラビリンチュラ類のうちヤブレツボカビ類とよばれる一群は、古くは卵菌に分類されていた[35])、ストラメノパイルの中でラビリンチュラ類はより初期に分岐しており、卵菌類と近縁ではない[3]。ストラメノパイルの共通祖先が紅藻との二次共生由来の葉緑体をもっており、偽菌類やラビリンチュラ類などの祖先で二次的に葉緑体・光合成能を失ったとする仮説(クロムアルベオラータ仮説)もあるが[43]、これを積極的に支持する証拠は見つかっていない[44]。
下位分類
卵菌は比較的大きなグループであり、2020年現在でおよそ100属1,700種ほどが知られている[17]。卵菌類は比較的古くから1つの綱、卵菌綱 (Oomycetes) にまとめられていた[23][35]。また、同じ範囲の生物群に対して、ツユカビ綱 (Peronosporomycetes) の名も提唱されている[2]。ただし、卵菌を1つの門、卵菌門 (Oomycota) として扱い、その一部のみに対してツユカビ綱の名を用いることもある[2][3](下記参照)。
菌体の体制や無性生殖様式、有性生殖様式、さらに20世紀末以降は分子形質に基づいて分類され、2023年現在では12目程度に分けられることが多い[3][17](下表3)。系統的には、これらの目は、多数の初期分岐群と2つの大きなグループ、ミズカビ群とツユカビ群に分けられる[2][3](下図10)。初期分岐群(basal oomycetes)はユーリカスマ目やフクロカビモドキ目など全て微細な内部寄生性種であり、特に2020年ごろから詳細に研究され多数知られるようになった[3]。ミズカビ群はフシミズカビ目(ときにアトキンシエラ目を分ける)とミズカビ目を、ツユカビ群はオオギミズカビ目、シロサビキン目、ツユカビ目を含む[3][17]。この2つの群は亜綱(ミズカビ亜綱、ツユカビ亜綱)に分類されることもあるが、卵菌類を門とし、この2群を綱(ミズカビ綱、ツユカビ綱(狭義))として扱うこともある[2][3]。ただし、これらの場合の初期分岐群の分類学的扱い(おそらく複数の亜綱または綱に分割する必要がある)は提唱されていない。ミズカビ群とツユカビ群は単系統群を形成すると考えられており、この単系統群は core oomycetes とよばれる[2][3]。
卵菌の初期分岐群の特徴から、卵菌の原始形質は海産で単純な全実性、内部寄生性であると考えられており、このような状態から、core oomycetes の共通祖先において淡水・陸上への進出、発達した菌糸体や腐生性の獲得が起こったとも考えられている[3]。ただし、core oomycetes の中にも海産、全実性、内部寄生性の種は存在し、このような方向への進化および逆方向への進化が何度も独立に起こったことが示唆されている[3]。また、有性生殖における配偶子嚢・配偶子の雌雄分化も core oomycetes に限られているが、同型配偶子嚢接合の種も知られており、このような雌雄分化はミズカビ群とツユカビ群で独立に起こった可能性も示唆されている[3]。ツユカビ群の中では、陸上植物絶対寄生性への進化が、シロサビキン目とツユカビ目において独立に起こったと考えられている[3]。
ツユカビ目は、植物絶対寄生性のツユカビ目(狭義)と腐生能をもつフハイカビ目に分けることもある[42][39]。しかし分子系統学的研究によって両群は連続的て明瞭に分けられないことが示されており、2023年現在ではふつうツユカビ目(広義)にまとめられている[3]。また、ツユカビ目の中でツユカビ科とフハイカビ科に分けられているが、分子系統学的研究に基づいて、後者から前者へ移された例(エキビョウキン属 Phytophthora など)も多い[3]。
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10. 卵菌綱の系統仮説の一例[3][15][16][17][45][46] |
表3. 卵菌の属までの分類体系の一例[4][47][35][42][48][49][50][17][15][46]
|
化石記録
卵菌類と考えられる化石は、古生代デボン紀前期のライニーチャートから報告されている[3]。また、石炭紀の地層からは、リンボク(小葉植物)やシダ植物、裸子植物に寄生していた可能性がある卵菌の卵胞子や生卵器の化石が報告されている[3][51][52][53]。また、白亜紀の琥珀からも卵菌類と考えられる構造が見つかっている[54]。
ギャラリー
-
 Albugo tragopogonis(シロサビキン目)が寄生したヒマワリ
Albugo tragopogonis(シロサビキン目)が寄生したヒマワリ -

-
 Phytophthora blight(ツユカビ目)が寄生したパパイヤ
Phytophthora blight(ツユカビ目)が寄生したパパイヤ -
 ジャガイモエキビョウキン(ツユカビ目)が寄生したジャガイモ
ジャガイモエキビョウキン(ツユカビ目)が寄生したジャガイモ -
 Peronospora manshurica(ツユカビ目)が寄生したダイズ
Peronospora manshurica(ツユカビ目)が寄生したダイズ -
 Hyaloperonospora(ツユカビ目)が寄生したヒメナズナ
Hyaloperonospora(ツユカビ目)が寄生したヒメナズナ -
 シロイヌナズナに寄生した Hyaloperonospora(ツユカビ目)の菌糸と植物細胞内に侵入させた吸器(トリパンブルー染色試料)
シロイヌナズナに寄生した Hyaloperonospora(ツユカビ目)の菌糸と植物細胞内に侵入させた吸器(トリパンブルー染色試料) -
 ツユカビ属(ツユカビ目)の菌糸体と気孔から伸ばした胞子嚢柄
ツユカビ属(ツユカビ目)の菌糸体と気孔から伸ばした胞子嚢柄 -
 Pseudoperonospora cubensis(ツユカビ目)の胞子嚢柄
Pseudoperonospora cubensis(ツユカビ目)の胞子嚢柄 -
 Pseudoperonospora(ツユカビ目)の遊走子嚢
Pseudoperonospora(ツユカビ目)の遊走子嚢 -
 エキビョウキン属(ツユカビ目)の卵胞子
エキビョウキン属(ツユカビ目)の卵胞子 -
 エキビョウキン属(ツユカビ目)の卵胞子
エキビョウキン属(ツユカビ目)の卵胞子


.jpg)






.jpg)


脚注
注釈
出典
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av Webster, J. & Weber, R. W. S. (2007). “Straminipila: Oomycota”. Introduction to Fungi 3rd Edition. Cambridge University Press. pp. 75–126. ISBN 978-0521014830
- ^ a b c d e f g h i j k l m n o p q r s t Beakes, G. W., Honda, D. & Thines, M. (2014). “Systematics of the Straminipila: Labyrinthulomycota, Hyphochytriomycota, and Oomycota”. In McLaughlin, D. J. & Spatafora, J. W.. THE MYCOTA, volume 7A. Systematics and Evolution Part A. Springer. pp. 39-97. doi:10.1007/978-3-642-55318-9_3
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf bg bh bi bj bk bl bm bn bo bp bq br bs bt bu bv bw bx by bz ca cb cc cd ce cf cg ch ci Beakes, G. W. & Thines, M. (2017). “Hyphochytriomycota and Oomycota”. In Archibald, J. M., Simpson, A. G. B. & Slamovits, C. H.. Handbook of the Protists. Springer. pp. 435-505. ISBN 978-3319281476
- ^ a b c d e f g h i j k l m n o 徳増征二 (2005). “卵菌門”. In 杉山純多. バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統. 裳華房. pp. 191–197. ISBN 978-4785358273
- ^ “The evolutionary phylogeny of Oomycetes—insights gained from studies of holocarpic parasites of algae and invertebrates”. 2012年10月25日閲覧。
- ^ Gordon W. Beakes & Sally L. Glockling & Satoshi Sekimoto (2012). “The evolutionary phylogeny of the oomycete “fungi””. Protoplasma 249: 3–19. doi:10.1007/s00709-011-0269-2.
- ^ Klinter, S., Bulone, V. & Arvestad, L. (2019). “Diversity and evolution of chitin synthases in oomycetes (Straminipila: Oomycota)”. Molecular Phylogenetics and Evolution 139: 106558. doi:10.1016/j.ympev.2019.106558.
- ^ 稲葉重樹 (2013). “1.8.6 サカゲツボカビ門”. In 日本菌学会 (編). 菌類の事典. 朝倉書店. pp. 41–42. ISBN 978-4254171471
- ^ Gachon, C. M., Strittmatter, M., Badis, Y., Fletcher, K. I., West, P. V. & Müller, D. G. (2017). “Pathogens of brown algae: culture studies of Anisolpidium ectocarpii and A. rosenvingei reveal that the Anisolpidiales are uniflagellated oomycetes”. European Journal of Phycology 52 (2): 133-148. doi:10.1080/09670262.2016.1252857.
- ^ Schnepf, E. & Drebes, G. (1977). “On the development of the marine, parasitic phycomycete Lagenisma coscinodisci (Lagenidiales)”. Helgoländer Wissenschaftliche Meeresuntersuchungen 29: 291-301. doi:10.1007/BF01614265.
- ^ 東條元昭 (2004). “ピシウム菌による病害”. 植物防疫 58 (3): 120–126.
- ^ 印東弘玄 (1985). “水棲菌類の観察方法”. 日本水処理生物学会誌 21 (1): 20-25. doi:10.2521/jswtb.21.20.
- ^ a b 稲葉重樹 (2014). “鞭毛菌類”. In 細矢剛, 国立科学博物館. 菌類のふしぎ 第2版. 東海大学出版部. pp. 20–28. ISBN 978-4486020264
- ^ a b Buaya, A. T., Ploch, S., Inaba, S., & Thines, M. (2019). “Holocarpic oomycete parasitoids of red algae are not Olpidiopsis”. Fungal Systematics and Evolution 4 (1): 21-31. doi:10.3114/fuse.2019.04.03.
- ^ a b c Buaya, A.T., Scholz, B. & Thines, M. (2021). “Taxonomy and phylogeny of Aphanomycopsis bacillariacearum, a holocarpic oomycete parasitoid of the freshwater diatom genus Pinnularia”. Mycol Progress 20: 289–298. doi:10.1007/s11557-021-01668-x.
- ^ a b c Buaya, A. T., Scholz, B. & Thines, M. (2021). “Sirolpidium bryopsidis, a parasite of green algae, is probably conspecific with Pontisma lagenidioides, a parasite of red algae”. Fungal Systematics and Evolution 7 (1): 223-231.. doi:10.3114/fuse.2021.07.11.
- ^ a b c d e f g Buaya, A. T. & Thines, M. (2020). “An overview on the biology and phylogeny of the early-diverging oomycetes”. Philippine Journal of Systematic Biology 14: 1-20. doi:10.26757/pjsb2020a14004.
- ^ Buaya, A. T., Ploch, S., & Thines, M. (2019). “Rediscovery and phylogenetic placement of Olpidiopsis gillii (de Wildeman) Friedmann, a holocarpic oomycete parasitoid of freshwater diatoms”. Mycoscience 60 (3): 141-146.
- ^ Buaya, A. T., & Thines, M. (2020). “Diatomophthoraceae–a new family of olpidiopsis-like diatom parasitoids largely unrelated to Ectrogella”. Fungal Systematics and Evolution 5 (1): 113-118.
- ^ a b Buaya, A. T. & Thines, M. (2020). “Bolbea parasitica gen. et sp. nov., a cultivable holocarpic parasitoid of the early-diverging Saprolegniomycetes”. Fungal Systematics and Evolution 6 (1): 129-137.
- ^ 竹中重仁 (2011). “菌寄生菌としての Pythium”. 植物防疫 65 (2): 117-120.
- ^ 築尾嘉章 & 東條元昭 (2013). “疫病菌とピシウム菌 類似点と相違点”. 植物防疫 67 (10): 536-552.
- ^ a b c ジョン・ウェブスター著 椿啓介、三浦宏一郎、山本昌木訳 (1985). “卵菌綱”. ウェブスター菌類概論. 講談社. pp. 139–187. ISBN 978-4061396098
- ^ Fontaine, M. C., Labbé, F., Dussert, Y., Delière, L., Richart-Cervera, S., Giraud, T. & Delmotte, F. (2021). “Europe as a bridgehead in the worldwide invasion history of grapevine downy mildew, Plasmopara viticola”. Current Biology 31 (10): 2155-2166. doi:10.1016/j.cub.2021.03.009.
- ^ Rizzo, D. M. & Garbelotto, M. (2003). “Sudden oak death: endangering California and Oregon forest ecosystems”. Frontiers in Ecology and the Environment 1 (4): 197-204. doi:10.1890/1540-9295(2003)001[0197:SODECA]2.0.CO;2.
- ^ a b 升屋勇人, 田端雅進, 市原優 & 景山幸二 (2019). “Phytophthora cinnamomi によるウルシ林の衰退 ―国産漆の新たなる脅威―”. 日本森林学会誌 101 (6): 318-321. doi:10.4005/jjfs.101.318.
- ^ 埋橋志穂美 (2011). “分子系統に基づく Pythium 属の新分類システム”. 植物防疫 65 (10): 587-592.
- ^ “日本植物病名データベース”. 農業生物資源ジーンバンク. 2023年9月12日閲覧。
- ^ 小林亨夫・勝本謙・安孫子和雄・阿部恭久・柿島真 (編), ed (1992). 植物病原菌類図説. 全国農村教育協会. pp. 58–71, 510–512. ISBN 9784881370452
- ^ Sekimoto, S., Yokoo, K., Kawamura, Y. & Honda, D. (2008). “Taxonomy, molecular phylogeny, and ultrastructural morphology of Olpidiopsis porphyrae sp. nov.(Oomycetes, straminipiles), a unicellular obligate endoparasite of Bangia and Porphyra spp.(Bangiales, Rhodophyta)”. Mycological Research 112 (3): 361-374. doi:10.1016/j.mycres.2007.11.002.
- ^ 東條元昭 (2009). “ノリアカグサレ菌の分類学的研究”. 日本水産学会誌 75 (5): 910-911. doi:10.2331/suisan.75.910.
- ^ 三浦正之, 畑井喜司雄, 東條元昭, 和田新平, 小林咲麗 & 岡崎巧 (2010). “アユ仔魚に発生した Pythium flevoense による内臓真菌症”. 魚病研究 45 (1): 24-30. doi:10.3147/jsfp.45.24.
- ^ a b c 上村兼輔 & 川井唯史 (2018). “新しい脅威である水カビ病について”. Cancer 27: 153-154. doi:10.18988/cancer.27.0_153.
- ^ Gaastra, W., Lipman, L. J., De Cock, A. W., Exel, T. K., Pegge, R. B., Scheurwater, J., ... & Mendoza, L. (2010). “Pythium insidiosum: an overview”. Veterinary Microbiology 146 (1-2): 1-16. doi:10.1016/j.vetmic.2010.07.019.
- ^ a b c d 井上浩, 岩槻邦男, 柏谷博之, 田村道夫, 堀田満, 三浦宏一郎 & 山岸高旺 (1983). 植物系統分類の基礎. 北隆館. p. 23–34
- ^ a b c 杉山純多 (2005). “菌類の多様性と分類体系”. In 杉山純多. バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統. 裳華房. pp. 30–55. ISBN 978-4785358273
- ^ a b c 稲葉重樹 (2014). “除外された偽菌類”. In 細矢剛, 国立科学博物館. 菌類のふしぎ 第2版. 東海大学出版部. pp. 99–105. ISBN 978-4486020264
- ^ Patterson, D.J. (1989). “Stramenopiles: chromophytes from a protistan perspective”. In Green, J.C., Leadbeater, B.S.C. & Diver, W.L.. Chromophyte Algae: Problems and Perspectives. Clarendon Press. pp. 357–379. ISBN 0198577133
- ^ a b Spring, O. (2012). “8. Oomycota”. In Frey, W. (eds.). Syllabus of Plant Families. A. Engler's Syllabus der Pflanzenfamilien Part 1/1. Borntraeger. pp. 98-105. ISBN 978-3-443-01061-4
- ^ Leonard, G., Labarre, A., Milner, D. S., Monier, A., Soanes, D., Wideman, J. G., ... & Richards, T. A. (2018). “Comparative genomic analysis of the ‘pseudofungus’ Hyphochytrium catenoides”. Open Biology 8 (1): 170184. doi:10.1098/rsob.170184.
- ^ a b c d Cho, A., Tikhonenkov, D. V., Hehenberger, E., Karnkowska, A., Mylnikov, A. P. & Keeling, P. J. (2022). “Monophyly of diverse Bigyromonadea and their impact on phylogenomic relationships within stramenopiles”. Molecular Phylogenetics and Evolution 171: 107468. doi:10.1016/j.ympev.2022.107468.
- ^ a b c 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一, ed (2013). “生物分類表”. 岩波 生物学辞典 第5版. 岩波書店. pp. 1653–1654. ISBN 978-4000803144
- ^ 矢吹彬憲・松本拓也 (2007). “クリプト藻類・ハプト藻類聞の姉妹群関係とクロムアルベオラータ仮説”. 藻類 55: 192-194.
- ^ Archibald, J. M. (2009). “The puzzle of plastid evolution”. Current Biology 19 (2): R81-R88. doi:10.1016/j.cub.2008.11.067.
- ^ Marco, T. & Young-Joon, C. (2016). “Evolution, Diversity, and Taxonomy of the Peronosporaceae, with Focus on the Genus Peronospora”. Phytopathology 106 (1): 6-18. doi:10.1094/PHYTO-05-15-0127-RVW.
- ^ a b Tsai, I. & Thines, M. (2023). “Adding a missing piece to the puzzle of oomycete phylogeny: the placement of Rhipidium interruptum (Rhipidiaceae)”. Fungal Systematics and Evolution 11 (1): 95-108. doi:10.3114/fuse.2023.11.08.
- ^ 杉山純多, ed (2005). “分類表 卵菌門”. バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統. 裳華房. pp. 397–398. ISBN 978-4785358273
- ^ “The MycoBank”. Robert, V., Stegehuis, G. & Stalpers, J.. 2023年8月12日閲覧。
- ^ Vilela, R., Humber, R. A., Taylor, J. W. & Mendoza, L. (2019). “Phylogenetic and physiological traits of oomycetes originally identified as Lagenidium giganteum from fly and mosquito larvae”. Mycologia 111 (3): 408-422. doi:10.1080/00275514.2019.1589316.
- ^ Duffy, M. A., James, T. Y. & Longworth, A. (2015). “Ecology, virulence, and phylogeny of Blastulidium paedophthorum, a widespread brood parasite of Daphnia spp.”. Applied and Environmental Microbiology 81 (16): 5486-5496. doi:10.1128/AEM.01369-15.
- ^ Dotzler, N., Krings, M., Agerer, R., Galtier, J. & Taylor, T. N. (2008). “Combresomyces cornifer gen. sp. nov., an endophytic peronosporomycete in Lepidodendron from the Carboniferous of central France”. Mycological Research 112 (9): 1107-1114. doi:10.1016/j.mycres.2008.03.003.
- ^ Harper, C. J., Krings, M., Galtier, J. & Taylor, T. N. (2016). “A microfossil with suggested affinities to the Peronosporomycetes Oomycota from the Carboniferous c. 330 Ma of France”. Nova Hedwig 103 (3-4): 315-326. doi:10.1127/nova_hedwigia/2016/0352.
- ^ Strullu-Derrien, C., Gèze, M., Spencer, A. R., De Franceschi, D., Kenrick, P., Selosse, M. A. & Knoll, A. H. (2021). “An expanded diversity of oomycetes in Carboniferous forests: Reinterpretation of Oochytrium lepidodendri (Renault 1894) from the Esnost chert, Massif Central, France”. Plos One 16 (3): e0247849.
- ^ “Introduction to the Oomycota”. 2012-09-閲覧。
外部リンク
- “The MycoBank”. Robert, V., Stegehuis, G. & Stalpers, J.. 2023年9月22日閲覧。 (英語)
- Fry, William E. & Grünwald, Niklaus J. (2010年). “Introduction to Oomycetes”. The American Phytopathological Society. doi:10.1094/PHI-I-2010-1207-01. 2023年9月23日閲覧。 (英語)