
「ワレイタムシ」の版間の差分
←新しいページ: 「{{生物分類表 | 色 = 動物界 | 名称 = '''ワレイタムシ''' | fossil_range = {{Fossil range|Late Silurian|Early Permian}} | 画像 = File:Eop…」 |
(相違点なし)
|
2020年11月5日 (木) 14:42時点における版
| ワレイタムシ | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ワレイタムシの1種 Eophrynus prestvicii の復元図
| |||||||||||||||||||||
| 保全状況評価 | |||||||||||||||||||||
| 絶滅(化石) | |||||||||||||||||||||
| 地質時代 | |||||||||||||||||||||
| シルル紀後期 - ペルム紀前期 | |||||||||||||||||||||
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 学名 | |||||||||||||||||||||
| Trigonotarbida Pocock, 1911 | |||||||||||||||||||||
| シノニム | |||||||||||||||||||||
|
Meridogastra Thorell & Lindström, 1885 Eurymarti Matthew, 1895 | |||||||||||||||||||||
| 和名 | |||||||||||||||||||||
| ワレイタムシ(割板虫、ワレイタムシ類) 割板類 | |||||||||||||||||||||
| 英名 | |||||||||||||||||||||
| Trigonotarbid | |||||||||||||||||||||
| 科 | |||||||||||||||||||||
ワレイタムシ(割板虫、割板類、ワレイタムシ類、Trigonotarbid、学名:Trigonotarbida[1][2])は、鋏角亜門・クモガタ綱のワレイタムシ目に分類される化石節足動物の総称である。クモらしい姿と横で数枚に分かれた背面の外骨格を特徴とする[3][4][1]。デボン紀と石炭紀の種類を中心とし、シルル紀からペルム紀にかけて生息していた[2]。
形態
-
 Archaeomartus roessleri の化石(背面)。横で3枚に分かれた一連の背板が見られる。
Archaeomartus roessleri の化石(背面)。横で3枚に分かれた一連の背板が見られる。 -


一見ではクモのような姿にあるが、体は丈夫で、前体と後体の間にあるくびれはクモほど顕著ではない。ナイフ状の鋏角や2対の書肺を有するなど、基本的体制は四肺類(クモ・ウデムシ・サソリモドキなど)のクモガタ類に共通する[1]が、横で3枚の外骨格に細分される背板などというクツコムシらしい特徴も見られる[5][4][1]。
前体
前体(頭胸部)は先節と直後6つの体節の癒合でできている合体節とされる[1]。背面は1枚の背甲(carapace または prosomal dorsal shield)に覆われ、その前半部は少なくとも1対の単眼である中眼(median ocelli)をもつ。一部の種では更に複数対のレンズでできている側眼(lateral eyes)が確認されており[6][3]、その数は最多15対で、複眼に近い形にある[7]。前体の腹面は前後で下唇と腹板があり、幅狭く、左右は脚の基節(最初の肢節)によって囲まれる[3][1]。
他のクモガタ類と同様、前体には鋏角1対・触肢1対・脚4対という計6対の付属肢(関節肢)がある[1]。鋏角は背甲の前部によって覆われ[8]、2節でできているナイフ型(亜鋏状)である[3][1]。鋏角全体は腹側に向かっており、左右は触肢の基節に挟まれる形で配置される[9]。基部の肢節は頑丈で内側に集密した剛毛があり、先端の肢節は牙のように尖って内側に曲がる[3][8]。ウデムシやサソリモドキのように牙の部分にも剛毛があるが、さほど集密ではない[9]。触肢は直後の脚より少し短い歩脚状で6節からなり、クモの触肢器やウデムシの棘のように顕著に特化した構造はないが、少なくとも一部の種はその先端に小さなはさみがある[4]。脚は歩脚状で7節からなり、先端は1対の爪がある[3][4][10]。4対の脚は通常ではほぼ同形だが、一部の種類は前方の脚がやや強大化している[10][11]。触肢と第1脚のそれぞれ基節は、口側に向かって配置される棘や剛毛などの突起物をもつ種類がある[12]。
上唇は鋏角と下唇の間に配置され、口は上唇と下唇の間のやや奥に開く。上唇と下唇の内側、いわゆる口への通り道には短い剛毛が走る[9]。
後体
後体(腹部)は楕円形で、腹面は椀のように膨らむのに対して背面は平たい[8][10]。12節の体節が含まれており[1]、そのほとんどが上下で背板と腹板によって表れる。第1節は少し幅狭くて知る限り腹板はなく[3][1]、その背板の前半部以上は直前の背甲の下に隠れ、凹凸で背甲と相互に嵌め込まれる構造(locking ridge)になる[5][4][13][1]。第2-3節の背板は往々にして前後癒合する[13][1]。本群の特徴でもあり、第2-9節のそれぞれの背板は横で3枚(中央1枚と左右2枚)の外骨格に細分される[1]。Anthracomartidae科の種類では更に細分化が進み、横で5枚になる[3][13]。最終の3節(第10-12節)の外骨格は環状にまとまり、背板と腹板の区別はない[1]。多くの場合、この3節は直前の第9節の腹面に配置されるため、背側からは観察できない[3][8]。中でも最終の2節は円筒状でとても小さく、まとめて後腹部(postabdomen)や尾部(pygidium)と呼ばれることもある[8][1]。尾節はない[1]。
古い文献では、第1節と背板の重なり具合・第2-3節の背板の癒合・最終2節の小ささなどの原因によって一部の体節が見逃され、後体の体節数は8から11節と見間違われることがある[8][1]。
四肺類と同様、2対の書肺は第2-3節の腹面に配置されるため、それを被った2枚の外骨格はおそらく腹板ではなく、付属肢由来の蓋板(operculum)であると考えられる[1]。蓋板の直後に「ventral sacs」という付属肢由来と思われる1対の突起物がある[8][1]。
| 後体の体節 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 構造 | 背板(locking ridge) | 背板(次の背板と癒合) 蓋板+書肺 |
背板(直前の背板と癒合) 蓋板+書肺 |
背板 腹板 ventral sacs |
背板 腹板 |
背板 腹板 |
背板 腹板 |
背板 腹板 |
背板 腹板 |
環状 | 環状(微小) | 環状(微小) |
他のクモガタ類との類似点
ナイフ型の鋏角や後体第2-3節にある2対の書肺を有するなど、クモガタ類の中でワレイタムシ類の基本体制は四肺類的である[1]。しかしその同時に、横で3枚に細分される背板・相互に嵌め込まれる背甲と第1背板(locking ridge)・先端に小さなはさみをもつ触肢など、クツコムシに共通する特徴もいくつか挙げられる[5][4][1]。その他にも、内側に剛毛のある上唇と下唇はコヨリムシ類に似通い[9]、蓋板の直後にventral sacを有することはウデムシ類に共通する[1]。
生態
ワレイタムシ類は陸棲で、書肺を通じて呼吸を行い、森林の地表を徘徊する捕食者であったと考えられる[9][8]。現生の四肺類のように、ナイフ型の鋏角で獲物を掴んでは消化液を吐き出して餌を体外消化し、上唇と下唇の隙間で液化した餌を吸収できたと考えられる[9]。口器(鋏角・上唇・下唇など)に配置される剛毛は現生の四肺類と同じく、液状の餌を濾過する機能をもつとされるが、ワレイタムシ類のそれが現生の四肺類に比べて特化は進んでいない(剛毛は短く少ない)ため、濾過効率は比較的に低かったと推測される[9]。多くの現生クモガタ類と同様、触肢と第1脚のそれぞれの基節に配置される突起物は、摂食行動を補助する口器であったと考えられる[12]。前方の脚を強大化した種類は、カニグモに似通う方法で捕食をしていたと推測される[11]。

.jpg)
Eophrynidae科のワレイタムシ類の後半部は、Laniatores亜目のザトウムシ類のように発達したコブや棘がある[11][8]。これらの特徴はそのザトウムシ類と同じく、天敵から身を守る役割を果したものであったと考えられており、収斂進化の例として挙げられる[8]。それに踏まえて、Eophrynidae科のワレイタムシ類は、Laniatores亜目のザトウムシ類を通じて広く見られるような警告色もあったのではないかという説も挙げられる[8]。
分類
| |||||||||||||||||||||||||||||||||||||||||||||
| Pantetrapulmonataの内部系統関係(†= 絶滅群、?= 不確実) |



.jpg)


クモ類・ウデムシ類・サソリモドキ類などを含んだ四肺類に共通する基本体制(主にナイフ型の鋏角と後体第2-3節にある2対の書肺)にあることから、クモガタ類の中で、ワレイタムシ類は少なくとも四肺類に近縁で、現生の四肺類に対して基盤的であることが広く認められる[6][9][14][15][3][4][8][16][1]。90前代前後の文献ではワレイタムシ類は四肺類に含まれた[9][14][15][3]が、2007年以降では四肺類の姉妹群として、共にPantetrapulmonataを構成するとされる方が一般的である[17][4][8][1][16]。
-

-
 クツコムシ類はワレイタムシ類のように横で3枚に分かれた背板をもつ。
クツコムシ類はワレイタムシ類のように横で3枚に分かれた背板をもつ。

_(cropped).jpg)
また、四肺類の体制をもつ上で前述のクツコムシ類らしい特徴(3枚に細分される背板・背甲と第1背板のlocking ridge・触肢先端の小さなはさみ)にもあることから、ワレイタムシ類は、系統的位置が不確実なクツコムシ類の類縁関係を示唆する存在ともされる[5][4]。もしクツコムシ類はワレイタムシ類の近縁であれば、この2群からなる系統群が四肺類の姉妹群になる[5][4][16]。広く認められる意見ではないが、クツコムシ類自体がワレイタムシ類の派生群ではないかという説もある[18][4]。
下位分類
2020年現在、9科36属70種のワレイタムシ類が記載される[2]。Jones et al. 2014 による系統解析では、Anthracomartidae科、Anthracosironidae科、Eophrynidae科は単系統群であるのに対して、Aphantomartidae科・Kreischeriidae科・Palaeocharinidae科・Trigonotarbidae科は非単系統群であると示唆される[13]。
(科は太字、1種のみを含む属は特記されない)
- (科)Anthracomartidae(石炭紀)
- (属)Anthracomartus(15種)
- (属)Brachypyge
- (属)Maiocercus
- (科)Anthracosironidae(デボン紀 - 石炭紀)
- (属)Anthracosiro(2種、石炭紀)
- (属)Arianrhoda(デボン紀)
- (属)Vratislavia(石炭紀)
- (科)Aphantomartidae(デボン紀 - ペルム紀)
- (属)Alkenia(デボン紀)
- (属)Aphantomartus(3種、石炭紀 - ペルム紀)
- (科)Archaeomartidae(デボン紀)
- (科)Eophrynidae(石炭紀)
- (属)Eophrynus(2種)
- (属)Nyranytarbus(2種)
- (属)Petrovicia
- (属)Planomartus
- (属)Pleophrynus(2種)
- (属)Pocononia
- (属)Somaspidion
- (属)Stenotrogulus
- (科)Kreischeriidae(石炭紀)
- (属)Anzinia
- (属)Gondwanarache
- (属)Hemikreischeria
- (属)Kreischeria
- (属)Pseudokreischeria
- (科)Lissomartidae(石炭紀)
- (属)Lissomartus(2種)
- (科)Palaeocharinidae(デボン紀)
- (属)Aculeatarbus
- (属)Gelasinotarbus(4種)
- (属)Gigantocharinus
- (属)Gilboarachne
- (属)Palaeocharinus(6種)
- (属)Spinocharinus
- (科)Trigonotarbidae(デボン紀 - 石炭紀)
- (属)Trigonotarbus(3種、デボン紀 - 石炭紀)
- (科未定)
- (属)Aenigmatarbus(石炭紀)
- (属)Anthracophrynus(石炭紀)
- (属)Aphantomartus(石炭紀)
- (属)Areomartus(石炭紀)
- (種)‘Eophrynus’ scharfi(ペルム紀)
- (属)Namurotarbus(石炭紀)
- (属)Palaeotarbus(シルル紀)
- (属)Permotarbus(ペルム紀)
- (属)Tynecotarbus(石炭紀)
ギャラリー
化石
-
 Archaeomartus roessleri
Archaeomartus roessleri -

-
 Cryptomartus rebskei
Cryptomartus rebskei -
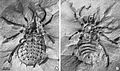 Eophrynus prestvicii
Eophrynus prestvicii -
 Eophrynus prestvicii
Eophrynus prestvicii -
 Kreischeria Vienna
Kreischeria Vienna -
 Permotarbus schuberti
Permotarbus schuberti





復元図
-
 Archaeomartus roessleri
Archaeomartus roessleri -
 Eophrynus prestvicii
Eophrynus prestvicii -
 Palaeocharinus rhyniensis
Palaeocharinus rhyniensis


脚注
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y Dunlop, Jason A.; Lamsdell, James C. (2017). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395. ISSN 1467-8039.
- ^ a b c Dunlop, J. A., Penney, D. & Jekel, D. 2020. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch, version 20.5
- ^ a b c d e f g h i j k Fayers, Stephen R.; Dunlop, Jason A.; Trewin, Nigel H. (2005-01-01). “A new early devonian trigonotarbid arachnid from the Windyfield Chert, Rhynie, Scotland”. Journal of Systematic Palaeontology 2 (4): 269–284. doi:10.1017/S147720190400149X. ISSN 1477-2019.
- ^ a b c d e f g h i j k Dunlop, J.; Kamenz, Carsten; Talarico, G. (2009). “A fossil trigonotarbid arachnid with a ricinuleid-like pedipalpal claw”. Zoomorphology. doi:10.1007/s00435-009-0090-z.
- ^ a b c d e Dunlop, J. A. (1996) (English). Evidence for a sister group relationship between Ricinulei and Trigonotarbida
- ^ a b Shear, William (1987) (英語). New Terrestrial Arachnids from the Devonian of Gilboa, New York (Arachnida, Trigonotarbida).
- ^ Miether, Sebastian T.; Dunlop, Jason A. (2016/07). “Lateral eye evolution in the arachnids”. Arachnology 17 (2): 103–119. doi:10.13156/arac.2006.17.2.103. ISSN 2050-9928.
- ^ a b c d e f g h i j k l m Dunlop, Jason; Garwood, Russell (2014). “Tomographic reconstruction of the exceptionally preserved trigonotarbid arachnid Eophrynus prestvicii”. Acta Palaeontologica Polonica. doi:10.4202/app.2012.0032.
- ^ a b c d e f g h i Dunlop, J. A. (Department of Geology (1994). “Filtration mechanisms in the mouthparts of tetrapulmonate arachnids (Trigonotarbida, Araneae, Amblypygi, Uropygi, Schizomida)” (English). Bulletin of the British Arachnological Society (United Kingdom).
- ^ a b c Garwood, Russell; Dunlop, Jason (2014-07). “The walking dead: Blender as a tool for paleontologists with a case study on extinct arachnids” (英語). Journal of Paleontology 88 (4): 735–746. doi:10.1666/13-088. ISSN 0022-3360.
- ^ a b c Garwood, Russell; Dunlop, Jason A.; Sutton, Mark D. (2009-12-23). “High-fidelity X-ray micro-tomography reconstruction of siderite-hosted Carboniferous arachnids”. Biology Letters 5 (6): 841–844. doi:10.1098/rsbl.2009.0464. ISSN 1744-9561. PMC 2828000. PMID 19656861.
- ^ a b Haug, Carolin (2020-08-13). “The evolution of feeding within Euchelicerata: data from the fossil groups Eurypterida and Trigonotarbida illustrate possible evolutionary pathways” (英語). PeerJ 8: e9696. doi:10.7717/peerj.9696. ISSN 2167-8359.
- ^ a b c d Jones, Fiona M.; Dunlop, Jason A.; Friedman, Matt; Garwood, Russell J. (2014-09-01). “Trigonotarbus johnsoni Pocock, 1911, revealed by X-ray computed tomography, with a cladistic analysis of the extinct trigonotarbid arachnids” (英語). Zoological Journal of the Linnean Society 172 (1): 49–70. doi:10.1111/zoj.12167. ISSN 0024-4082.
- ^ a b Dunlop, Jason A. (1996-03-01). “A redescription of the trigonotarbid arachnid Pocononia whitei (Ewing 1930)” (英語). Paläontologische Zeitschrift 70 (1): 145. doi:10.1007/BF02988273.
- ^ a b Rößler, Ronny; Dunlop, Jason A. (1997-09). “Redescription of the largest trigonotarbid arachnid —Kreischeria wiedei Geinitz 1882 from the Upper Carboniferous of Zwickau, Germany” (英語). Paläontologische Zeitschrift 71 (3-4): 237–245. doi:10.1007/BF02988493. ISSN 0031-0220.
- ^ a b c Garwood, Russell J.; Dunlop, Jason (2014-11-13). “Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders” (英語). PeerJ 2: e641. doi:10.7717/peerj.641. ISSN 2167-8359.
- ^ Shultz, Jeffrey W. (2007-06). “A phylogenetic analysis of the arachnid orders based on morphological characters” (英語). Zoological Journal of the Linnean Society 150 (2): 221–265. doi:10.1111/j.1096-3642.2007.00284.x. ISSN 1096-3642.
- ^ Karsch, F. (1892). “Ueber Cryptostemma Guér. als einzigen recenten Auslaüfer der fossilen Arachnoideen-Ordnung der Meridogastra Thor” (英語). Berliner entomologische Zeitschrift 37 (1): 25–32. doi:10.1002/mmnd.18920370108.