
頭足類の体

頭足類の体(とうそくるいのからだ)では、軟体動物門 頭足綱 Cephalopodaに属する生物の体制について解説する。
頭足類は、オウムガイ亜綱 Nautiloidea、および絶滅したアンモナイト亜綱 Ammonoidea、そしてオウムガイ類以外の現生の頭足類を含む鞘形亜綱 Coleoideaに大別される。現生の鞘形類は十腕形上目 Decapodiformes(イカ類)と八腕形上目 Octopodiformes(広義のタコ類)に二分され、十腕形類にはトグロコウイカ目 Spirulida、コウイカ目 Sepiida(ダンゴイカ目 Sepiolidaを分離することもある)およびツツイカ目 Teuthida(開眼目 Oegopsida及び閉眼目 Myopsidaの2群に分けることも多い)が、八腕形類にはタコ目(八腕目)Octopodaとコウモリダコ目 Vampyromorphaが含まれる。鞘形類にはオーラコケラス目 Aulacocerida、ベレムノイド目 Belemnitida、フラグモテウティス目 Phragmoteuthidaといった化石頭足類も含まれる。
頭足類は無脊椎動物の中で最も賢く、最も複雑であるとされる[1]。大きな脳や発達した視覚と筋肉質の腕により、複雑な行動を進化させ、好奇心と明確な個性をもつ[1]。
体制


頭足類は軟体動物の中では大型である[2]。体は左右相称で前後方向に伸長し[2]、腹側に深い外套腔が形成される[3]。体は主に前方 (anterior)の足と頭部が一体化した頭足塊(とうそくかい、head-foot)と後方 (posterior)の内臓塊(ないぞうかい、visceral mass)からなる[3][4]。
頭足塊には腕と呼ばれる足があり、これがこの群、頭足類 Cephalopodaの名称の由来となっている。腕は口を取り囲むように生え、各腕に列をなした吸盤をそなえる[2]。口にはよく発達した口球があり、その中に嘴状の顎板と歯舌をそなえる[2]。頭部の両側によく発達した1対の眼があり、鞘形類ではレンズをそなえた精巧な眼をもつ[2]。
後方に突出した内臓塊は外套膜により包みこまれ細長い胴部を形成する[2]。外套腔は前方に向かって開き、その腹側の縁には筒状に突出した漏斗がある[2]。外套腔内には鞘形類では1対、オウムガイ類は2対の鰓をそなえる[2]。軟体動物は普通貝殻をもつが、頭足類では貝殻は退化的で現生のものは一般に内在性[2]。オウムガイ類は螺旋状に巻いた大きな外在性の貝殻をもち、その中に内臓塊が収まる[2]。コウイカ類では背部の外套膜の中に甲と呼ばれる石灰質の内在性の貝殻が残っているが、タコ類などでは貝殻は完全に退化する[2]。貝殻は多室構造をもち、この構造はオウムガイ類の外在性の殻だけでなくイカ類の内在性の殻にも認められる[2]。鞘形類には墨汁腺をもつものがあり、直腸付近に開口する[2]。中腸腺はよく発達し、肝臓域と膵臓域が分化している[2]。循環系は閉鎖血管系で、鞘形類では通常の心臓のほかに、鰓の基部に1対の鰓心臓をもつ[2]。腎臓は1対[2]。生殖巣は対をなさず、いずれも外に開口する[2]。発達した神経節が頭部に集中して高度な中枢神経系(脳)をなす[2]。
感覚器官
頭部には眼があり、その後腹側には微小な嗅覚器官(きゅうかくきかん、olfactory organ、または嗅孔 olfactory pit[5])が開孔する[4]。眼の前方には涙孔(るいこう、lacrymal pore)が開孔する[5]。頭蓋軟骨を取り出して後側から見ると腹側に平衡胞(へいこうほう、statocyst)が透けて見える[4]。
眼
眼 (eye)は頭部の側面の左右に1対あり[4]、よく発達する[3][6]。オウムガイ類はレンズのないピンホール型の杯眼を持つ[3][7]。鞘形類はレンズを伴う高度に発達した眼を持つ[3]。
オウムガイ類の杯眼は腹足類のもつ杯眼のより発達した型で、感覚上皮が体表から陥入し、通光性の分泌物で満たされ、内腔が小孔により外界に通じる[7]。ピンホールカメラの機能があると考えられている[7]。
鞘形類のカメラ眼では水晶体(レンズ)、虹彩、毛様体、眼瞼(がんけん、eyelid, orbit[5])など脊椎動物と類似の構造が作られ形態視 (form vision)機能が発達する[7]。頭足類の眼は脊椎動物の眼に酷似するが、頭足類では視神経が網膜の外側から伸び盲点がないため、内側から伸びた視神経が束になって網膜を貫通し、盲点のできる脊椎動物の眼と区別される[3]。つまり発生学的にも対光性網膜を持つ杯眼としての基本的性格が保たれていると解釈される[7]。コウイカ類の眼の表面は角膜 (cornea)に覆われ閉じている[4]。角膜を切り開くと、レンズ (lens)と網膜 (retina)が見える[4]。レンズは上下が毛様体により吊られており、網膜はその奥にある[6]。スルメイカの網膜には光を受容する桿状細胞が160,000個/mm2並んでいる[6][8]。レンズの上下には虹彩 (iris)があり、光量を調節する[4]。ツツイカ類は眼の特徴により、透明な膜で眼を覆われる閉眼類 Myopsidaと膜を持たない開眼類 Oegopsidaに二分される[6]。また、少なくとも数種は色を識別できることが解っている[9]。
口
口(くち、mouth)の周辺は唇(くちびる、lip)で囲まれ、その周辺は囲口膜(いこうまく、buccal membrane)に覆われる[5]。口より腹側には受精托(じゅせいたく、semibal receptacle)がある[5]。
口球
脳と口の間には口球(こうきゅう、buccal mass, buccal bulb)があり、その先端には顎板が飛び出す[4]。口球は軟体動物における消化管先端の膨らんだ筋肉質の塊状部分で、口腔を囲み歯舌、舌軟骨、顎板を納める[10]。歯舌嚢が付随し、唾腺が開口する[10]。脳神経節の前方に口球神経節(こうきゅうしんけいせつ、buccal ganglion)があり、頭足類では上下2節に分かれている[10]。
顎板
.jpg)
.jpg)
顎板(がくばん、beak, jaw plate)は口球中に見られるクチクラ質(キチン質)の嘴状物 (rostrum)である[11]。通称「カラストンビ」と呼ばれる[4][11]。上下1対あり、下側の方が強く突出する[4]。上顎板(じょうがくばん、upper jaw plate[12])は細身でカーブが緩いためカラスに、下顎板(かがくばん、lower jaw plate[12])は強く曲がり、猛禽類の嘴のようなのでトンビに喩えられる[13]。歯舌前端に対面する口腔の背面に位置し、食物を噛み切るとともに上顎部が歯舌で傷つくのを防いでいる[11]。捕食者であるクジラや鳥に飲み込まれてもイカの体で最後まで消化されないのは顎板のみであり[13]、食性研究者は捕食者の胃内に残された顎板を手がかりに、その捕食者はどのような頭足類を餌としているかを追求する[14]。
歯舌

歯舌(しぜつ、radula)は口球中に見られるクチクラ質の基底膜上に多数の小歯(しょうし、tooth)が無数の横列をなして並ぶ鑢様のリボンである[15]。小歯の先端には鋭い歯尖(しせん、cusp)があり、小歯には3種類ある[15]。1個の中心歯(ちゅうしんし、中歯、central tooth, rhachidian tooth)の両脇に1ないし数個の側歯(そくし、lateral teeth)、更にその外側に数個の縁歯(えんし、marginal teeth)が並んでいる[15]。
神経系
脳
中枢神経系は多くの軟体動物では発達せず、食道周辺にある3対前後の神経節が横連合と縦連合により環状に結び付けられ、食道神経環となる[4]。しかし頭足類ではその中枢神経系が著しく発達し、脳 (brain)を形成する[4]。脳は左右眼の中間に位置し、眼の基部には視葉(しよう、optic lobe)が発達する[4]。硬い頭蓋軟骨に覆われ、脳の中央部には食道が貫通する[4]。脳は大きく、他の軟体動物における脳神経節だけでなく側神経節、足神経節、内臓神経節とも相同だと考えられている[9]。
星状神経節
漏斗軟骨器付近の外套膜内面には頭足類に特有の星状神経節(せいじょうしんけいせつ、stellate ganglion)が見られる[4]。多方向に分岐しており、外套膜に神経を送っている[4]。
腕

本群における腕(わん、うで、arm)は、他の動物における足に該当する器官である。これは動物学的には足であるが、餌を捕らえたり、雌を抱きかかえたり、物を運ぶ機能を持つため、慣習的に「腕」と呼ぶ[8]。
オウムガイ類では腕は触手(しょくしゅ、cirrus)とも呼ばれ、数十本の短い触手が2列になって口の周囲を取り囲む[2](全部で約90本[3])。
鞘形亜綱では、背側から腹側に向かい、左右それぞれ第1腕 (I, first arm, dorsal arm)、第2腕 (II, second arm)、第3腕 (III, third arm)、第4腕 (IV, fourth arm, ventral arm)の4対の腕が口を取り囲むようにあり、更に十腕形上目では第3腕と第4腕の間から触腕と呼ばれる特殊な腕が伸びる[8][3]。この2本の触腕の有無により八腕形類と十腕形類に分けられる[3]。これには例外があり、十腕形類にも拘らずヤツデイカ科 Octopoteuthidaeに属する種およびテカギイカ科 Gonatidaeのタコイカ Gonatopsis borealisの成体は腕を8本しか持たない[3][8]。幼若個体には10本腕があるが、成長に伴い触腕が2本とも失われる[3][8]。
種によっては腕の間に傘膜(さんまく、umbrella)または腕間膜(わんかんまく、interbrachial membrane, interbrachial web[5])と呼ばれる膜が発達する[3]。
雄では一部の腕に生殖のための特殊化が見られ、交接腕となる[3]。
鞘形類の腕には吸盤 (sucker)や鉤(かぎ、hook)がある[3]。オウムガイ類の触手には吸盤がなく、粘着性の分泌物で付着する[3]。
交接腕
交接腕 (こうせつわん、ヘクトコチルス、hectocotylus, hectocotylized arm)は雄が持つ、雌に精包およびその中の精子を渡すために変形した腕である[16]。化茎腕や生殖腕とも呼ばれる[16]。成熟とともに一定の腕が交接腕に変形する現象を化茎現象(かけいげんしょう、hectocotylization)[16]または交接腕化という。交接腕はその種によって変化の様式と腕の位置が決まっている[16]。イカでは腕の末端よりの吸盤のいくつかが消失し、吸盤柄が櫛の歯状に並ぶものや特別な膜や突起が生じるものがある[16]。タコでは腕の先端が匙状になり、そこまで精包が通る溝が走るものがある[16]。普通コウイカ類のもつ交接腕は再利用できるが、アオイガイ科やアミダコ科の雄では一生に一度しか交接できず、精子を満載した交接腕が雌の体内に挿入されると、切り離され雌の外套腔内に残る[16][17]。英名 hectocotylusの名は、ギリシア語で100を意味するἑκατόν (hekatón)に由来する接頭辞 hecto-と小さな器を意味するギリシア語のκοτύλη (kotýlē)からなり[18]、1829年、ジョルジュ・キュヴィエがアオイガイの交接腕を寄生虫と誤認し百疣虫 Hectocotylus octopodis Cuvier, 1829と命名したことによる[16][17]。
オウムガイ類の交接腕はスペイディクス (spadix)とよばれ、勃起して精子を雌に渡す役割を持っている[17]。
-
 タコの交接腕の模式図
タコの交接腕の模式図 -
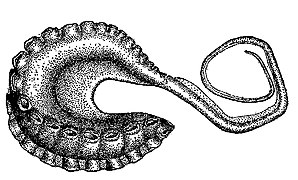


吸盤
_on_tentacular_club.jpg)
吸盤(きゅうばん、sucker)は腕および触腕の口側 (oral)に一定に配列し、他物に吸着するための盤状構造[19]。イカ類(十腕形類)の吸盤は基部が柄のように細くなっており、吸盤の内部には角質環(かくしつかん、chitinous ring, horny ring)と呼ばれる硬い有機質のリングがある[3]。一般に4縦列 (quadserial)で配置される[19]。タコ類には柄も角質環もなく、この吸盤の構造の違いが「イカ」と「タコ」を区別する最も重要な形質である[3][19]。メンダコ類の腕には吸盤に加えて触毛(しょくもう、cirrus)が生えている[3]。タコ類の吸盤は吸着によって、イカ類の吸盤は角質環でしがみつくことによって機能する[3]。
触腕

触腕 (tentacle)は十腕形類のみがもつ、伸縮自在で、餌を捕獲するたの特殊な腕である[8]。触腕は先端部が広くなり、その部分だけに吸盤があるが[4]、この部分を触腕掌部[5](しょくわんしょうぶ、tentacle club)または触腕頭[12](腕頭部[20])という。またそれを繋ぐ伸縮自在の部分を触腕柄(柄部[20])と呼ぶ。眼の下にあり第3腕と第4腕の間に開口するポケット中に触腕は折りたたまれる[4]。コウイカ類の触腕は完全にポケットの中に収めることができ、外からは見えないこともある[4]。ツツイカ類では完全には収納できず、常時出ているが、腕と同長程度に収縮させている[20]。
開眼類では触腕の吸盤に機能により形態分化が見られ、基部 (carpus)、掌部 (manus)および先端部 (dactylus)の3群に分かれる[20]。
漏斗
頭足類は優れた運動性能を持つが、それは腹側にある漏斗(ろうと、funnel)から水を吹き出しジェット推進(jet propulsion)により移動することによる[3]。漏斗は外套膜縁と頭部の間からのぞいている[4]。漏斗の向きは自在に動かすことができ、これによって遊泳方向を調節できる[3]。漏斗はオウムガイ類では閉じた管ではないが、鞘形類では完全に閉じた水管状となる[3]。漏斗の内部には弁状の構造、漏斗弁(ろうとべん、funnel valve)があり、これで水流が逆流しないよう調節する[4]。また漏斗内部には背側漏斗器(はいそくろうとき、dorsal funnel organ)および腹側漏斗器(ふくそくろうとき、ventral funnel organ)がある[5]。
漏斗の周辺の頭部の最後端部は頸部(けいぶ、neck)と呼ばれる[3]。漏斗の左右には頸部と外套膜を咬み合わせる漏斗軟骨器(ろうとなんこつき、funnel locking cartilage, funnel cartilage)と呼ばれる軟骨の塊が1対あり、外套膜の外套軟骨器(がいとうなんこつき、mantle cartilage[12])と嵌合する[3][4]。漏斗軟骨器は釦穴(ぼたんあな、button-hole)、外套軟骨器は釦(ぼたん、button)とよばれることもある。[12]。漏斗軟骨器の後方には1対の太い漏斗牽引筋(ろうとけんいんきん、funnel retractor muscle、漏斗下制筋 funnel depressor muscleとも[12])が伸び、殻に付着する[4]。
外套膜

頭足類の体は外套膜(がいとうまく、mantle)により覆われる[4]。外套膜が貝殻に付着する二枚貝類や貝殻と離れる腹足類と違い、現生頭足類の多くは貝殻が外套膜中に内在する[4]。外套膜の内側には外套腔(がいとうこう、pallial cavity[4], mantle cavity[2])と呼ばれる空所が形成され、呼吸器官や排出器官が集中する[4]。外套腔中には肛門、腎臓の排出孔(腎門)、生殖門が開口する[4]。
頭足類の外套膜には環状筋と放射筋が重なり合っている[3]。環状筋が緩み、放射筋が収縮すると頭と外套膜の間から水が取り入れられ、次に環状筋が収縮すると襟が頭部に押しつけられ、水は漏斗から排出される[3]。
貝殻
_001.jpeg)

頭足類には貝殻 (shell)をもつ種ともたない種があり、その殻も外在性のものと内在性のものがある[3]。頭足類の貝殻は外殻性から内殻性に進化したと考えられており、現生種では内殻性が一般的である[3]。
オウムガイ類は外殻性の貝殻を持ち、内部は隔壁(かくへき、septum)によって仕切られる[3]。隔壁をもつ部分は気房(air chamber)または房錐(phragmocone)、隔壁の内部は気室(きしつ、chamber, camera)と呼ばれる[3]。隔壁の内部は浮力調節器官として機能する連室細管(れんしつさいかん、siphunclular tube)によって貫かれ、その内部には体管索(たいかんさく、siphuncular cord)と呼ばれる組織に満たされている[3]。動物体は殻の末端部にある住房(じゅうぼう、body chamber, living chamber)に入っており、動物体は成長するにつれて定期的に前方へ移動し、後方に新しい隔壁をつくる[3]。現生オウムガイ類の場合隔壁は30以上形成され、殻の殻頂部には胚の段階で形成される胚殻(はいかく、embryonic shell)が残される[3]。
鞘形類では内殻性の殻をもつが分類群によって退化、消失する[3]。トグロコウイカ Spirula spirulaではオウムガイ類と同様の螺旋状に巻いた殻を持つが、殻は内殻性で大部分が外套膜によって包まれている[3]。コウイカ科 Sepiidaeの貝殻は背腹に押しつぶされたような形の甲となる[3]。
ツツイカ類では貝殻は有機質の膜になり、軟甲と呼ばれる[3]。
タコ類の成体では貝殻は形成されない[3]。ただしアオイガイ科 Argonautidaeのアオイガイ Argonauta argoやタコブネ Argonauta ahiansも螺旋状に巻いた外在性の貝殻をもつが、この貝殻の内部には隔壁がなく、卵を保護するために雌の第1腕から二次的に分泌される[2][3]。他の頭足類の殻は腕ではなく外套膜から分泌されるため、両者の殻は起源が異なる[2][3]。
甲
甲(こう、cuttlebone, sepion)は、(狭義の)コウイカ類がもつ内在性の貝殻である[21]。オウムガイ類やトグロコウイカの形態とは一見無関係だが、その断面は内部には薄い板状の隔壁が密集しており、その間の隙間がオウムガイ類やトグロコウイカの気室に相当する[3]。
コウイカ属 Sepiaでは甲の後端には棘(きょく、spine[5])または嘴部(しぶ、rostrum[12])をもつ。
軟甲

軟甲(なんこう、gladius, pen)はツツイカ類の甲で、さらに退化して石灰質を失い、殻皮質 (コンキオリン、conchiolin)のみとなったもの[22]。
鰭
十腕類は外套膜に鰭(ひれ、fin)をもつ。ツツイカ類では外套膜の後方に菱形につくが、コウイカ類では外套膜の側縁全長に亘っている[23]。前者のタイプを俗に「耳」、後者のタイプを「えんぺら」と呼ぶ[23]。またミミイカの鰭は半円状で、後方で連続しない[24]。
タコ類は普通鰭を持たないが、深海付近に生息するヒゲダコ科やメンダコ科のタコは鰭がある[25]。
鰓
軟体動物の鰓は櫛鰓(くしえら、しっさい、ctenidium, comb gill)または本鰓と呼ばれ、1対の血管を含む鰓軸(さいじく、ctenidial axis)の両側に多数の櫛歯状に平行して鰓葉(さいよう、ctenidial lamella)が並ぶ[4][26]。鰓軸中の血管は入鰓血管(にゅうさいけっかん、afferent branchial vessel)と出鰓血管(しゅっさいけっかん、efferent branchial vessel)とよばれる[26]。オウムガイ類及びアンモナイト類は2対の鰓を持つのに対し、鞘形類は1対の鰓を持ち[26]、それぞれ別名四鰓亜綱 Tetrabranchiaと二鰓亜綱 Dibranchiaと呼ばれる[27]。
消化器官
コウイカ Sepia esculentaの消化管は口球の後方に、食道、胃、胃盲嚢、消化腺、腸、肛門と続く[4]。食道 (esophagus)は脳を貫通し、内臓の中央部まで伸びる[4]。胃 (stomach)は大きく膨らみ、胃の左側に胃本体よりも大きい胃盲嚢(いもうのう、caecum)がある[4]。胃盲嚢はほぼ一巻きの螺旋を描く[4]。消化腺 (digestive gland)は胃よりも前にあり、1対の左右相称である[4]。消化腺と胃は細長い管で連結し、管周辺には消化腺付属体 (digestive gland appendage, pancreatic appendage)が発達する[4]。腸 (intestine)は単純で細く、1度捩れた後に前方に伸び、外套膜内に開口する肛門 (anus)で終わる[4]。
頭足類の食道は膨張部を持つことがあり、嗉嚢(そのう、crop)とよばれる[9]。ここにはかなり大きな食物を一時的に収容することができ、食物を磨り潰す筋肉質の胃に送る[9]。消化は細胞外で行われ、胃から始まるが、その殆どは胃盲嚢か胃盲嚢と消化腺の両方で行われる[9]。水溶性の栄養分はこの2器官で行われる[9]。胃盲嚢の繊毛運動で内容物を攪拌し、粘液と混ぜ合わされ、残渣は胃盲嚢と胃の結合部からつながる腸に送られる[9]。腸は短く、消化管の末端部分は直腸 (rectum)と呼ばれる[9]。
墨汁嚢
墨汁嚢(ぼくじゅうのう、inc sac)は頭足類に特有の墨汁を溜めておく嚢状器官[28]。直腸の背側にあり、タコ類では肝臓に埋没している[28]。墨汁嚢の後端にある墨汁腺(ぼくじゅうせん、ink gland)から墨汁が分泌され、墨汁管 (ink sac duct)を通って前側に送られる[28][4]。墨汁管は肛門付近で腸と連結し、肛門を通じて放出される[4]。
墨汁 (sepia, ink)はセピオメラニンやプテリンなどを含む[28]。イカの墨汁は捕食者の目を欺くダミー効果で、タコでは煙膜効果があると言われている[28]。深海性のものは墨汁嚢を持たない[28]。
循環器系
頭足類の血管系は閉鎖血管系で、よく発達する[4]。コウイカの心臓は2心房1心室である[4]。心臓から出る大動脈は、長太く頭部に達する前大動脈と内臓の後部に達する後大動脈に分かれる[4]。動脈より腹側には頭部から戻る血液が流れる大静脈は消化腺直下の正中線上にあり、その後側では左右に分岐する[4]。分岐の先は腎嚢付属体に包まれ、鰓心臓に接続する[4]。出鰓血管はすぐに心房につながる[4]。
鰓心臓
鰓心臓(えらしんぞう、branchial heart)は入鰓血管の基部にあり、律動的に収縮する膨大部である[29]。入鰓血管中に静脈血を送り込んでいる[29]。櫛鰓の数に応じ、鞘形類(二鰓類)では合計2個ある[29]。壁は腺様組織からなり、他の軟体動物の囲心腺と相同な腺様の付属体がある[29]。十腕類では鰓心臓と付属体はともに体腔中にあるが、八腕類では付属腺だけが体腔中にある[29]。
排出器官
頭足類の排出器官は腎嚢(じんのう、renal sac)と呼ばれる淡褐色で背腹方向に折れ曲がった1対または2対の大きな薄壁の嚢である[30]。内端の腎囲心管は囲心腔に、外端の腎門(じんもん、renal opening, nephridial apperture[12]、尿乳頭 urinary papilla[12]、または排出孔 excretory pore[4])は外套腔に開口する[30]。入鰓血管の壁にある腺様組織(腎嚢付属体、renal appendage[4])及び鰓心臓の腺様付属体(鰓心臓付属体 branchial heart appengdageの中にある囲心嚢付属体 pericardial appendage[4])も排出機能を持ち、この排出物も腎嚢内に送り出される[30][4]。内腔は真体腔の一部で、腎腔(じんこう、urocoel)と呼ばれる[30]。腎嚢中には二胚動物が多数生息する[30]。
生殖器官
頭足類は雌雄異体で、生殖器官は雌雄で異なる[4]。生殖器官は生殖巣(せいしょくそう、gonad)と生殖輸管(せいしょくゆかん、gonoduct)からなり、頭足類の生殖輸管は左側にしかない[4]。開口部は生殖門(せいしょくもん、genital opening)と呼ばれる[12]。
雌性生殖器
雌は大きな塊状の卵を卵巣 (ovary)内にもつ[4]。体の左側だけに単純な輸卵管(ゆらんかん、oviduct)があり、前側に伸びて外套腔に開口する[4]。輸卵管と離れた肛門付近には大きな腺状組織があり、後方の1対は卵包腺(らんぽうせん、nidamental gland、または卵嚢腺[12]、纏卵腺[12])とよばれ、その前方に付属卵包腺(ふぞくらんぽうせん、accessory nidamental gland、または副卵嚢腺[12])がある[4]。
雄性生殖器
雄は無構造の白い塊状の精巣(せいそう、testis)をもつ[4]。体の左側には輸精管(ゆせいかん、vas deferens、または精管[12])があり、細く折れ曲がって塊状となる[4]。精巣からつながった輸精管は貯精嚢(ちょせいのう、seminal vesicle、Vesicula seminalis)、副精莢腺(ふくせいきょうせん、accesory spermatophoric gland、または摂護腺 proostate gland)、精莢管(せいきょうかん、spermatophoric duct)、精莢嚢(せいきょうのう、spermatophoric sac)、陰茎(いんけい、penis)と続き、生殖門に至る[12]。雄の精子は精莢(せいきょう、spermatophore)[12]や精包(せいほう)[4]と呼ばれるものに入れて雌に渡されるが、輸精管の一部に精包を分泌するための精包腺(せいほうせん、spermatophoric gland)があり、完成すると精莢嚢[12](精包嚢またはニーダム嚢 Needam's sacとも[4])に貯蔵される[4]。
脚注
- ^ a b Kröger, Vinther & Fuchs (2011)
- ^ a b c d e f g h i j k l m n o p q r s t u v 上島励 (2000)
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao 佐々木猛智 (2002)
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf 佐々木 (2008)
- ^ a b c d e f g h i 奥谷喬司, 田川勝 & 堀川博史 (1987), pp. 27–28
- ^ a b c d 奥谷喬司 (2009), pp. 43–47
- ^ a b c d e 巌佐庸 et al. (2013), p. 1373
- ^ a b c d e f 奥谷喬司 (2010), pp. 2–34
- ^ a b c d e f g h Kozloff (1990), pp. 447–462
- ^ a b c 巌佐庸 et al. (2013), p. 436
- ^ a b c 巌佐庸 et al. (2013), p. 207
- ^ a b c d e f g h i j k l m n o p q 広島大学生物学会 (1971), pp. 79–85, plate 79–85
- ^ a b 奥谷喬司 (2009), pp. 48–49
- ^ 窪寺恒己 (2014年). “頭足類の顎板による種査定に関するマニュアル”. 2019年12月11日閲覧。
- ^ a b c 巌佐庸 et al. (2013), p. 585
- ^ a b c d e f g h 巌佐庸 et al. (2013), p. 452
- ^ a b c Staaf (2018), pp. 100–103
- ^ Webster (1958), p. 840
- ^ a b c 巌佐庸 et al. (2013), p. 313
- ^ a b c d 巌佐庸 et al. (2013), p. 679
- ^ 瀧巖 1999, p. 366.
- ^ 瀧巖 1999, p. 333.
- ^ a b 奥谷喬司 (2009), pp. 40–43
- ^ 土屋光太郎, 山本典瑛 & 阿部秀樹 (2002), p. 39
- ^ 土屋光太郎, 山本典瑛 & 阿部秀樹 (2002), p. 119
- ^ a b c 巌佐庸 et al. (2013), p. 346
- ^ 巌佐庸 et al. (2013), p. 988
- ^ a b c d e f 巌佐庸 et al. (2013), p. 1304
- ^ a b c d e 巌佐庸 et al. (2013), p. 146
- ^ a b c d e 巌佐庸 et al. (2013), p. 712
参考文献
- Kröger, Björn; Vinther, Jakob; Fuchs, Dirk (2011-08). “Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules: Extant cephalopods are younger than previously realised and were under major selection to become agile, shell-less predators” (英語). BioEssays 33 (8): 602–613. doi:10.1002/bies.201100001.
- Kozloff, Eugene N. (1990). Invertebrates. Saunders College Pub
- Webster, Noah (1958). Webster's New Twentieth Century Dictionary of the English Language, Unabridged (Second ed.). The World Publishing Company. pp. 100, 840, 1296. OCLC 1020875243
- 巌佐庸、倉谷滋、斎藤成也、塚谷裕一『岩波生物学辞典 第5版』岩波書店、2013年2月26日。ISBN 9784000803144。
- 上島励 著「21章 軟体動物門」、岩槻邦男 監修、馬渡峻輔 監修、白山義久 編『無脊椎動物の多様性と系統』裳華房〈バイオディバーシティ・シリーズ〉、2000年11月30日、169-188頁。ISBN 4785358289。
- 奥谷喬司、田川勝、堀川博史『日本陸棚周辺の頭足類 大陸棚斜面未利用資源精密調査』社団法人 日本水産資源保護協会、1987年、36–39頁。OCLC 18206695。
- 奥谷喬司『イカはしゃべるし、空も飛ぶ』講談社〈ブルーバックス〉、2009年8月20日。ISBN 978-4-06-257650-5。
- 奥谷喬司『日本のタコ学』東海大学出版会、2013年6月1日、2–27頁。ISBN 9784486019411。
- 奥谷喬司『新鮮イカ学』東海大学出版会、2010年7月20日。ISBN 9784486018759。
- 佐々木猛智「頭足類(Cephalopoda)」『貝の博物誌』東京大学総合研究博物館、2002年。
- 佐々木, 猛智「軟体動物の解剖 : コウイカ・サザエ・ホタテガイ」『化石』第84巻、日本古生物学会、2008年、86–95頁、doi:10.14825/kaseki.84.0_86。
- Staaf, Danna 著、日向やよい 訳、和仁良二 監修 編『Squid Empire: The Rise and Fall of the Cephalopods [イカ4億年の生存戦略]』エクスナレッジ、2018年、100–102頁。ISBN 978-4-7678-2499-4。OCLC 1043342704。
- 土屋光太郎、山本典瑛、阿部秀樹『イカ・タコガイドブック』TBSブリタニカ、2002年4月27日、39, 119頁。ISBN 4-484-02403-9。
- 広島大学生物学会 著、池田嘉平、稲葉明彦 編『日本動物解剖図説』森北出版、1971年9月22日。ISBN 4-627-26020-2。
- 瀧巖『動物系統分類学5(上)』中山書店、1999年。