
胚発生

胚発生(はいはっせい、英語: embryogenesis)または生物学における発生(はっせい)とは、多細胞生物が受精卵(単為発生の場合もある)から成体になるまでの過程を指す。広義には老化や再生も含まれる。発生生物学において研究がなされる。
前成説と後成説[編集]
古くは、卵または精子の中に小さな人の形をしたもの(これをホムンクルスという)があらかじめ存在し(つまり、受精卵の中に、子孫の雛形がある)、発生はホムンクルスが大きくなる過程であるという前成説が唱えられた時代があった。後の研究で、そのようなものは存在しないことが明らかとなった。他方、受精卵の中には雛形に相当するものは何もなく、次第に形ができて来るという考え方を後成説という。
実際には顕微鏡を使用して、細胞レベルの観察が行われるようになって、具体的な発生の過程が観察できるようになった(もっとも、研究の初期には「顕微鏡を通してホムンクルスが観察された」といったような報告がされたこともあった)。動物の発生については多くの研究がなされているが、植物についてはかなり遅れて研究がなされた。
動物の発生過程[編集]
様々な無脊椎動物の発生過程の研究から、動物の発生には、一定の共通する型があると考えられるようになった。

前核の出現[編集]
受精が成立すると、前核(prenuclear; PN)が出現する。精子の進入後約6時間で見え始め、約20時間で見えなくなる。雄性前核は受精卵で形成される精子由来の核、雌性前核は卵子由来の核である。雄性前核と雌性前核の融合により、父親と母親の遺伝情報を保持する子供の核が作られる。
体外受精では受精操作の翌日、PNの観察により受精の確認を行う。2PNは正常受精卵。1PNは2PNが融合後消える途中の場合と異常受精の場合がある。3PN以上は2つ以上の精子が入ってしまった異常受精である。0PNはすでに前核が消失してしまったか未受精を示す。
卵割[編集]
多細胞動物の発生は、受精卵の細胞分裂、いわゆる卵割から始まる。卵割は同調的な分裂により、細胞数を2の級数で増やす。通常は2細胞期に左右に、4細胞期に前後に、8細胞期に腹背に分かれる。卵黄の多い卵では、このとき動物極側(卵黄が少ない)の方が細胞が小さくなる。

胞胚期[編集]
ある程度細胞数が増えると、多くの場合、内部に空洞ができる。その外側は一層の細胞に覆われた形になる。この時期を胞胚(ほうはい)期と呼ぶ。胞胚の内部の空洞は卵割腔(らんかつこう)または、胞胚腔(ほうはいこう)と呼ばれる。ウニ卵は胞胚期に孵化し、表面に繊毛を持って泳ぐ。
卵割腔がなく、内部まで細胞で満たされるものもある。また、脊椎動物では複数の細胞層が生じる。

原腸の形成[編集]
胞胚の表面の細胞層が内部に入り込む。これは陥入(かんにゅう)と呼ばれる。そして、一つの口を持った袋を内部に形成する。これが消化管の始まりである。この袋は原腸と呼ばれ、その出入り口は原口と呼ばれる。この時期の胚を嚢胚(のうはい)または原腸胚(げんちょうはい)とよび、その形成を原腸胚形成 (Gastrulation) という。
刺胞動物や扁形動物では消化管には一つしか出入り口がない。その他の動物では消化管は管状である。そのような動物では、原口の反対側に新たにもう一つ出入り口ができる。このとき、どちらが口になるかは動物門によって異なる。軟体動物、節足動物、環形動物など、多くの無脊椎動物など原口動物(先口動物ともいう)では原口が口になるが、棘皮動物や脊椎動物など新口動物(後口動物ともいう)では原口は肛門になる。
胚葉の分化[編集]
原腸が陥入したことで、それまで平等に並んでいた細胞が、内側と外側に分かれたことになる。そこで、外に残った細胞群を外胚葉(がいはいよう)、内側に入った細胞群を内胚葉(ないはいよう)と呼ぶ。外胚葉からは主に表皮と神経が、内胚葉からは消化管が形成される。刺胞動物は、このような構造をほとんどそのままに成体になるので、二胚葉性動物と言われる。
それ以外の動物では、外胚葉と内胚葉の隙間に細胞群が入り込み、そこで発達を始める。この細胞群を中胚葉(ちゅうはいよう)とよぶ。中胚葉からは筋肉、血管系などが作られる。また、中胚葉からは体腔が作られる。これらの動物は三胚葉に分類される。
形態形成[編集]
ここから後は各器官が形成される段階になる。これを形態形成という。
形成体[編集]
ドイツのハンス・シュペーマンはイモリ胚での移植実験(1924年)から、原口背唇部(げんこうはいしんぶ)に分化を引き起こす作用を発見し、原口背唇部を形成体(オーガナイザー)と名付け、未分化の細胞群に分化を促す形成体の作用を誘導と呼んだ。
また、ドイツのヴァルタ―・フォークトが、イモリの胚を部分的に染色する「局所生体染色法」を開発した。フォークトはこれにより染色された胚がどのように分化するかの追跡調査を行い、胚が将来形成する原基の位置を示した原基分布図(予定運命図)を作成した(1929年)。
-
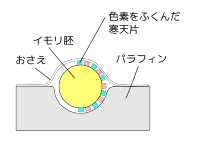 よくある、局所生体染色法の図。
よくある、局所生体染色法の図。 -
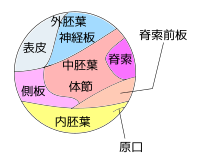 よくある、イモリの原基分布図。
よくある、イモリの原基分布図。


シュペーマンの実験において、原口背唇部の誘導の後に次々と組織・器官が形成されたことから、誘導の連鎖が推測された。
誘導の連鎖[編集]
一例として目の形成過程を以下に示す。
- 胚の原口背唇†(一次形成体)
- 脳の両側に形成される眼胞(がんぽう)→眼杯(がんぱい。二次形成体)
- 水晶体(三次形成体)
- 表皮の表層に働きかけ角膜を誘導する。
- † 原口背唇とは原口の上下にある細胞群のうち、背側(動物極側)にあるものをいう。原口を口とすると、この部位は唇の位置に当たるため、このような名前がつく。動物極とは、卵ができる過程で極体が放出される側を指す(動物極の反対側は植物極と呼ばれる)。極体とは、細胞核を持つが細胞質を持たない細胞で、減数分裂の途中で放出されるが、後々に退化して消えてしまう細胞。
誘導のメカニズム[編集]
誘導は分泌性因子を介した細胞間相互作用により行われると考えられるが、しばしば多数の因子が複雑な制御系を形成しており、分子メカニズムが解明されていないことも多い。例えば、上記の形成体の誘導にはWntシグナル系と呼ばれるシグナル伝達系路が重要であることが知られているが、その際にWntシグナル系がどのようにして活性化されるのかについては不明な点も多い。また、眼胞/眼杯による誘導には神経成長因子および骨形成因子が関与していると考えられているが、その詳細は明らかではない。
関連項目[編集]
参考文献[編集]
- 鈴木孝仁; 本川達雄『新生物II』(新課程)数研出版〈チャート式シリーズ〉、2005年11月1日。ISBN 4-410-11881-1。
- 『編・太田次郎 本川達雄 高等学校新編:生物I 新興出版社啓林館 平成17年12月10日発行 平成14年3月10日検定済・高等学校理科用』
外部リンク[編集]
- “発生と腫瘍形成の共通性”. 理研CDB - 科学ニュース. 独立行政法人理化学研究所 (2007年11月1日). 2010年12月19日閲覧。
この項目は、生物学に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:生命科学/Portal:生物学)。 |