
ミズクラゲ
| ミズクラゲ | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ミズクラゲの成体
| |||||||||||||||||||||
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 学名 | |||||||||||||||||||||
| w:Aurelia coerulea | |||||||||||||||||||||
| 和名 | |||||||||||||||||||||
| ミズクラゲ(水海月 | |||||||||||||||||||||
| 英名 | |||||||||||||||||||||
| (Asian) Moon Jelly |
ミズクラゲ(学名Aurelia coerulea)は、ミズクラゲ科に属するクラゲの一種。
日本近海でも最も普通に観察できるクラゲである。
形態
[編集]成体で傘の直径15 - 30cm、それ以上のものも稀に見られる。傘には、縁辺部に中空の細く短い触手が一列に無数に並ぶ。傘の下側の中央に十字形に口が開き、その4隅が伸びて、葉脈の位置で二つ折りにしたヤナギの葉のような形の4本の口腕となる。

体は四放射相称で、口腕の伸びる方向を正軸、その中間の軸を間軸という。間軸の方向に4つの丸い胃腔があり、馬蹄形の生殖腺に取り囲まれる。このため、4つの眼があるように見える。まれに五放射、六放射になっているものも見られるが、基本的な体の作りは同じである。
-
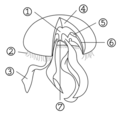 1.胃腔 2.触手 3.口腕 4.中膠 5.生殖腺 6.放射管 7.口
1.胃腔 2.触手 3.口腕 4.中膠 5.生殖腺 6.放射管 7.口

生態
[編集]成体は通年出現するが、特に夏に多く、また秋にも小さなピークがあることが知られている[1][2]
時間帯によって生息水深を変えていることが報告されている。福井県の夏の観察では夜はやや深い所に多いという[3]。明暗および水温に左右されているものと見られ、発電所からの温排水によって表層が30度を超すと浮上しなかった。塩分濃度の影響については研究者によって見解が分かれる。季節性もあり、晩春は朝夕に表層に出現し、夜は低層にいるという[4]。長崎県では、秋の個体は時間帯に関わらず低層に留まることが多かった[5]
塩分濃度の影響も指摘されており、特に大量の真水が流入し表層に真水層が形成されると水深以外にも様々な影響があるという[6]
餌となるのは主に動物性プランクトンで、時に仔魚を捕食する。遊泳運動は捕食活動も兼ねており、傘を開閉することで縁辺部の触手の間で海水が濾過され、そこに浮遊する動物プランクトンが触手に捕らえられる。この時クラゲが起こす水流より、遊泳能力が大きいプランクトンは脱出できるが、遊泳能力の低いものは捕獲されやすいことが示唆されている[7]。貝の幼生も捕食しており[8]、ミズクラゲが大発生するとアサリなどの漁獲に影響があると見られている。また、他のクラゲを捕食する事例も観察されている。
餌は触手の刺胞に刺されて麻痺すると同時に粘着性の刺糸に絡めとられ、粘液と繊毛運動により、傘の縁、縁弁の中央の8(胃腔の数の2倍)箇所に団子状に集められる。間欠的に口腕が触手をなでるときに口腕の溝の内側に餌が包み込まれ、繊毛運動によって口に運ばれる。胃腔消化された餌の粒子や液は、放射管から水管(血管のようなもの)を通って全身へと運ばれる。呼吸においても、同じ器官を通して体全体に拡散された海水より酸素を取り込む。
傘を開閉する運動は遊泳と捕食のためだけのものではなく、循環機能を働かせるための運動でもあり、つまり体そのものが心臓の役割を果たしている。また、クラゲ類は中枢神経系を欠き、体のどの部分にあっても一つの神経細胞が命令を下すと、新幹線並の速度で神経伝達が行われ、相対的に統合した運動を行うことができる。傘の縁の8カ所に、光の明暗を感じる眼点とバランスを取るための平衡器を備えた感覚器がある。
天敵は多く、クラゲ食のクラゲ、ウミガメ、マンボウ[9]などがいる。
福井県での観察では産卵、および発生の初期はアマモやホンダワラのような海藻(海草)が繁茂する湾の奥の浅い場所ではないかとみられている[10][11]。
遊泳能力は高くないと見られているが、ミズクラゲが集まるところは塩素濃度が高いという報告がある[12]
生活環
[編集]
成熟した雄から精子が水中に放たれ、雌がそれを取り込んで受精をする。受精卵は雌の口腕の保育嚢に運ばれ、卵割を繰り返して体表に繊毛が生じてプラヌラ幼生にまで成長してから海中に泳ぎ出る。プラヌラの長さは約0.2mmで体表の繊毛を動かして回転しながら遊泳する。数日間遊泳した後、適当な付着基盤に付着して変態を開始する。先端に触手が伸び、定着してから15時間ほどでイソギンチャクに似た、ポリプと呼ばれる段階に成長する。変態直後のポリプは2本の触手を持ち、中央に口が開く。触手には刺胞が備わっており、餌を摂ることができる。ポリプは摂餌により成長し、触手の数は4、8、16本と増え、24本に達することもある。成長したポリプは無性生殖で増殖し、コロニーを形成する。無性生殖は直接、あるいは走根(ストロン)上に新しいポリプを形成する出芽と、体が水平方向にのびて、縦に2つに分かれる分裂が主であるが、ポリプが移動した後にシストを作ることもある。
ポリプは非常に優れた再生能力を持つ。例えば細かくすり潰してしまっても、しばらくするとバラバラになった細胞組織が集まり始め、最終的にポリプを再生する。
東京湾での観察ではポリプの死亡率は高く、付着性の貝類との競争などが指摘されている[2]。福井県での観察では、産卵は早春から初夏にかけて行われると推測されている[13]。
成長していったポリプは徐々に体にくびれ(環溝)ができ始め(横分体形成、ストロビレーション)、くびれはさらに発達して8枚の縁弁が形成される。この時期のポリプを横分体(ストロビラ)と呼ぶ。横分体はくびれを増やしながら伸びてゆき、やがて先端の触手は吸収されて消失し、各節が分離して海中へと泳ぎ出す。この3mmほどの花のような形をしたエフィラはその一つ一つがミズクラゲの幼生である。腕状の縁弁の間が成長して円形になった時点で成体とほぼ同じ形の稚クラゲ(直径1から2cm)になる。エフィラと稚クラゲの間にメテフィラと呼ばれる段階を区別することもある。
プラヌラ→ポリプ→ストロビラ→エフィラ→(メテフィラ)→稚クラゲ→成体
一部の地域個体群では、プラヌラ幼生が直接エフィラ幼生に変態してそのままクラゲになるものがあることが知られている。
分布
[編集]環太平洋地域。西は日本や韓国からアメリカ西海岸、オーストラリア一帯まで広く分布する。かつては全世界的に分布するとされたが、後述のように近年分類が細分化しており、日本近海でみられるのはAurelia coeruleaとされている。
人間との関係
[編集]刺胞毒
[編集]刺胞を持つが、ヒトに対する毒性は低く遊泳中に皮膚の角質の薄い顔面にふれたときに、人によっては多少の痛みを感じる程度である。ザリガニに対する毒性試験で猛毒のハブクラゲの1⁄4程度の毒を持っているとされ、分子量43000の酸性タンパク質が毒性物質の主成分と考えられている[14]。繁殖力が強く、生活環も明らかにされていることから研究用途に使われることが多い。また、その半透明の美しい姿は人々を魅了し、ペットとしてもよく飼育されている。
漁業・工業被害
[編集]漁網に大量に入ると引き上げ時に網を損傷させたり、捕獲した魚介類が毒や酸欠により死亡、またクラゲ混入により冷却能力が低下し鮮度が落ちる[15]ことなどが問題となる。また、工場や発電所の取水口を詰まらせ、冷却水不足によるトラブルを引き起こすこともあり対策が求められている[16]。
取水口のクラゲ対策としては、網を設置したり、エアーポンプによるクラゲの強制浮上、ポンプによる汲み上げ、水流発生装置などがあるという[17]。
廃棄物として処理する際には水揚げ後いくらか乾燥させた後に、埋めることが多い。漁業の場合は船上で破砕処理できるクラゲカッターも導入されている。放線菌から採取されたタンパク質分解酵素にクラゲを漬け、50℃で処理すると数十分で完全に液化するという[18]。肥料としての利用も検討されている。クラゲが含む窒素はアンモニア態窒素が多いという[19]。
食用・薬用
[編集]クラゲ類を喫食したことによる食物アレルギーが報告されている[20][21]が、ミズクラゲでは今のところ知られていない。
クラゲに含まれるムチンからさらに高度のムチンを合成する原料にできるのではと注目されている[22]。
養殖魚の飼料として利用すると横臥状態を改善できるという報告がある[23]。また、ミズクラゲ溶解物は養殖業に壊滅的な被害を与える赤潮を引き起こす渦鞭毛藻に対して殺藻効果があるという[24]
分類学上の位置づけ
[編集]従来学名には Aurelia aurita が当てられてきたが、形態的に酷似する未知の隠蔽種が複数存在する可能性が指摘されていた[25]。
2017年、遺伝子及び形態的な分析の結果、A. auritaは大西洋沿岸に限定して分布する種で、太平洋に分布する種は別種 A. coeruleaとすることが妥当とされた。この種の模式標本の採取地はオーストラリアとされている。この論文内ではインド洋種と地中海種についてもそれぞれ別種扱いが妥当とされている[26]。本項ではこの説に従い、日本産ミズクラゲをAurelia coeruleaとして扱う。
名前
[編集]和名ミズクラゲは、体が透けており水と見分けがつきにくいこと、また水分を多く含み水っぽいことなどからの命名と見られる。傘に4つの目玉状の模様が現れることから、ヨツメクラゲ(四ツ目クラゲ)と呼ぶ地方がある。これは目ではなく胃腔と生殖腺である。
英名は他の近縁種も含めてmoon jelly(月のクラゲ)と呼ばれる。傘の丸い形を満月に、および目玉模様をクレーターに例えたものとみられる。特に本種を指定したいときはAsian(アジアの)など分布地を付けて呼ぶ。
脚注
[編集]- ^ 安田徹 (1969) 福井県浦底湾におけるミズクラゲの生態-I -成体の出現状況-. 日本水産学会誌 35(1), p.1-6. doi:10.2331/suisan.35.1
- ^ a b 石井晴人, 渡邊朋子 (2002) ミズクラゲのポリプ期における生態. Sessile Organisms 19(2), p.121-128. doi:10.4282/sosj.19.121
- ^ 安田徹 (1975) 福井県浦底湾におけるミズクラゲの生熊-XII. 水産増殖 22(3-4), p.128-134. doi:10.11233/aquaculturesci1953.22.128
- ^ 安田徹 (1973) 福井県浦底湾におけるミズクラゲの生態-IX -1970年春期における成体型の昼夜移動-. 日本水産学会誌 39(11), p.1145-1150. doi:10.2331/suisan.39.1145
- ^ 松下吉樹, 鈴木秀規, 梶川和武 (2011) 超小型データロガーを用いたミズクラゲの鉛直移動追跡. 水産工学 47(3), p.197-206. doi:10.18903/fisheng.47.3_197
- ^ 野村英明, 石丸隆 (1998) 東京湾におけるクラゲ類(刺胞動物及び有櫛動物)の最近15年間の出現状況. 海の研究 7(2), p.99-104. doi:10.5928/kaiyou.7.99
- ^ Costello, J.H., Colin, S.P. (1994) Morphology, fluid motion and predation by the scyphomedusa Aurelia aurita . Marine Biology 121, 327–334. doi:10.1007/BF00346741
- ^ 高橋宏司, 澤田英樹, 益田玲爾 (2019) アサリ浮遊幼生および着底直後稚貝の潜在的捕食者の探査. 水産増殖 67(3), p.275-279. doi:10.11233/aquaculturesci.67.275
- ^ 澤井悦郎, 安部奏, 横地和正, 前里馨 (2021) 横浜・八景島シーパラダイスの半野生環境下で撮影された水面でミズクラゲを捕食するマンボウ. Ichthy, Natural History of Fishes of Japan 13, p.13-17. doi:10.34583/ichthy.13.0_13
- ^ 安田徹 (1981) ミズクラゲの発生場所について. 付着生物研究 3(1), p.1-8. doi:10.4282/sosj1979.3.1
- ^ 安田徹 (1969) 福井県丹生浦湾におけるミズクラゲのエフイラと小型成体の出現状況. 水産増殖 17(1), p.33-39. doi:10.11233/aquaculturesci1953.17.33
- ^ 桑原連, 佐藤修一, 野口信彦 (1969) ミズクラゲの生態学的研究-I. 日本水産学会誌 35(2), p.156-162. doi:10.2331/suisan.35.156
- ^ 安田徹 (1971) 福井県浦底湾におけるミズクラゲの生態-IV -傘径組成の月変化と繁殖期について-. 日本水産学会誌 37(5), p.364-370. doi:10.2331/suisan.37.364
- ^ 永井宏史 (2005) クラゲ類の大量発生とそれらを巡る生態学・生化学・利用学 III-2. 刺胞毒とその利用. 日本水産学会誌 71(6), p.989-990. doi:10.2331/suisan.71.989
- ^ 林陽子 (1998) 定置網漁船の魚倉内における海水氷冷却に及ぼすミズクラゲの阻害作用. 日本水産学会誌 64(6), p.1046-1052. doi:10.2331/suisan.64.1046
- ^ 磯舜也 (1971) 海水の直接利用における取水設備について. 日本海水学会誌 24(4), p.139-148. doi:10.11457/swsj1965.24.139
- ^ 伊与久智明, 松浦正己, 熊本均 (2003) クラゲ排除装置について. Techno marine 日本造船学会誌 872,p.193-203. doi:10.14856/technom.872.0_193
- ^ 土井宏育, 武田美貴雄, 岡達三, 野々村禎昭 (2006) 微生物酵素によるミズクラゲの分解処理. 日本海水学会誌 60(6), p.426-433. doi:10.11457/swsj1965.60.426
- ^ 福士惠一, 有田翼, 南伸哉, 北方基一, 辻本淳一, 山下雄 (2006) クラゲに含まれる窒素およびリンの形態変化とクラゲ肥料化法. 日本海水学会誌 60(3), p.173-180. doi:10.11457/swsj1965.60.173
- ^ 鈴木慎太郎ら (2017) クラゲによるアレルギーの1成人例. アレルギー 66(6), p.804-808. doi:10.15036/arerugi.66.804
- ^ 窪田祥平ら (2017) タコクラゲ摂取によりアナフィラキシーを起こした14歳男児例. アレルギー 66(6), p.809-812. doi:10.15036/arerugi.66.809
- ^ 丑田公規 (2010) クラゲから抽出した新しいムチンの応用の可能性. 化学と生物 48(1), p.6-7. doi:10.1271/kagakutoseibutsu.48.6
- ^ 宮島(多賀)悠子, 益田玲爾, 栗原紋子, 山下洋, 竹内俊郎 (2014) ミズクラゲ給餌によるマダイ人工孵化稚魚の横臥傾向の改善. 日本水産学会誌 80(6), p.934-945. doi:10.2331/suisan.80.934
- ^ 半田慎也, 広海十朗, 内田直行 (1998) ミズクラゲ自己溶解液の新型赤潮鞭毛藻 Heterocapsa circularisquamaに対する殺藻効果. 日本水産学会誌 64(1), p.123-124. doi:10.2331/suisan.64.123
- ^ Dawson M.N. (2003) Macro-morphological variation among cryptic species of the moon jellyfish, Aurelia (Cnidaria: Scyphozoa). Marine Biology 143: 369-379. doi:10.1007/s00227-003-1070-3
- ^ Simonetta Scorrano, Giorgio Aglieri, Ferdinando Boero, Michael N. Dawson, Stefano Piraino (2017) Unmasking Aurelia species in the Mediterranean Sea: an integrative morphometric and molecular approach. Zoological Journal of the Linnean Society 180(2): 243-267. doi:10.1111/zoj.12494
参考文献
[編集]- 内田亨、時岡隆・谷田専治 著「腔腸動物」、内田亨・監 編『動物系統分類学2 中生動物 海綿動物 腔腸動物 有櫛動物』 2巻、中山書店、1961年。OCLC 672633613。
- 坂田明『クラゲの正体』晶文社、1994年。ISBN 4794961855。OCLC 47346957。
- 安田徹『ミズクラゲの研究』日本水産資源保護協会、1988年。NAID 10007462948。
- 岩間靖典 著、江ノ島水族館・監 編『クラゲ-その魅力と飼い方』誠文堂新光社、2001年。ISBN 4416701284。OCLC 54730374。
- 安田徹、上野俊士郎・足立文 著、安田徹・編 編『海のUFOクラゲー発生・生態・対策』恒星社厚生閣、2003年。ISBN 4769909764。OCLC 167761178。
- ジェーフィッシュ 著、久保田信・上野俊士郎・監修 編『クラゲのふしぎ』技術評論社、2006年。ISBN 4774128570。OCLC 675346795。
外部リンク
[編集]- 日本プランクトン学会
- 標本・資料統合データベース > 動物研究部 > 刺胞・有櫛動物 国立科学博物館
- Animal Diversity Web > Aurelia aurita (英語) 形態、生態など解説
| 典拠管理データベース: 国立図書館 |
|---|
この項目は、生物学に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:生命科学/Portal:生物学)。 |