
「筋細胞」の版間の差分
Smilesworth (会話 | 投稿記録) en:Muscle cell (21:17, 18 October 2021 UTC) の翻訳をもとに作成。長大なEvolution節は割愛 |
(相違点なし)
|
2021年11月16日 (火) 15:17時点における版
筋細胞(きんさいぼう、英: muscle cell, myocyte)は筋肉を構成する細胞であり、筋芽細胞と呼ばれる胚性前駆細胞から発生する[1]。Myocyteという語は心筋細胞または平滑筋細胞を指す場合に用いられ、これらはどちらも小細胞である[2]。骨格筋細胞は長く線維状で多くの核を持ち、筋線維と呼ばれる[2]。
筋芽細胞は筋形成の過程で融合して合胞体と呼ばれる多核細胞となり、骨格筋を形成する[3][4]。骨格筋細胞と心筋細胞は筋原線維とサルコメアを持ち、横紋筋組織を形成する[5]。
心筋細胞は心腔の壁面に心筋を形成し、細胞の中心に1つの核を持つ[6]。心筋細胞は隣接する細胞と介在板によって連結されており、目に見える単位で連結されているものは心筋線維(cardiac muscle fiber)と呼ばれる[7]。
平滑筋細胞は食道や胃の蠕動運動などの不随意運動を制御する。平滑筋には筋原線維やサルコメアがないため、横紋筋ではない。平滑筋細胞は1つの核を持つ。
構造
筋細胞の微視的構造は他の細胞とは大きく異なるため、特有の用語が用いられている。筋細胞の細胞質は筋形質(sarcoplasm)、滑面小胞体は筋小胞体(sarcoplasmic reticulum)、細胞膜は筋鞘(sarcolemma)と呼ばれている[8]。筋鞘は刺激を受け取り伝導する。
骨格筋細胞

骨格筋細胞は筋肉内で収縮する個々の細胞であり、長い線維状の外見をしているため一般的には筋線維として知られている[9]。一例として、青年男性の上腕二頭筋には約253,000個の筋線維が含まれている[10]。骨格筋線維は多核化している唯一の筋細胞であり、その核は筋核(myonucleus)と呼ばれる。この多核化は筋形成の過程で筋芽細胞が融合することで生じたものであり、各筋芽細胞が新たに形成された筋細胞または筋管(myotube)に核を提供する[11]。融合はMyomakerやMyomergerと呼ばれる筋特異的タンパク質に依存している[12]。
横紋筋にはミオフィラメントの長いタンパク質鎖から構成される筋原線維(myofibril)が含まれている。ミオフィラメントにはthin(細い)、thick(太い)、elasticの3つの種類が存在し、協働して筋収縮を生み出す[13]。細いフィラメントは主にアクチン、太いフィラメントは主にミオシンからなり、互いに滑ることで筋収縮時に線維の長さを短くしている。3番目の種類のelastic filamentは非常に巨大なタンパク質チチンから構成される。
横紋筋の縞模様は、ミオシンが暗く見えるフィラメントを形成してA帯を構成し、アクチンが明るく見えるフィラメントを形成してI帯を構成することで作られている。筋線維内での収縮の最小単位はサルコメアと呼ばれ、2つのZ線(Z板、Z膜)の間の反復単位である。また筋形質には、運動量が増えたときに細胞にエネルギーを供給するグリコーゲンや、筋活動に必要となるまで酸素を貯蔵する赤色色素であるミオグロビンが含まれている[13]。
筋小胞体は平滑小胞体が特殊化したものであり、筋線維の各筋原線維の周囲にネットワークを形成している。このネットワークは、2つの終末槽(terminal cisterna)と呼ばれる嚢状構造と、細胞を貫通する1本のT管(T-tubule、横行小管)からなるグループによって構成されている。これら3つの構成要素は筋小胞体のネットワーク内で三連構造(triad)となって存在しており、各T管の両側に2つの終末槽が位置している。筋小胞体はカルシウムイオンの貯蔵庫の役割を果たしており、T管を活動電位が広がると、筋小胞体へシグナル送られて膜チャネルからカルシウムイオンが放出され、筋収縮が刺激されることとなる[13][14]。
骨格筋の各筋線維の末端では、筋小胞体の外層が筋腱接合部(myotendinous junction)で腱線維と結合する[15][16]。筋線維内では核は筋鞘に押されて平坦な形状になっている。筋線維の多核状態は発生学的には、複数の筋芽細胞が融合して筋線維を形成する際に各筋芽細胞が1つの核を持ち込んだものである[13]。
心筋細胞
心筋細胞の細胞膜には、介在板やT管などの特殊な領域が存在する。細胞膜は約50 nmの幅の薄層で覆われている。この薄層は、基底板(lamina densa)と透明帯(lamina lucida)の2層に分けられる。この2層の間にはカルシウムイオンなどいくつかのイオン種が存在する[17]。
心筋は骨格筋と同様に横紋筋であり、骨格筋細胞と同様に筋原線維、ミオフィラメント、サルコメアが存在する。細胞膜は直径約10 nmのanchor fiberによって細胞骨格に固定されている。Anchor fiberは主にZ線に位置して溝を形成し、T管が通る。そのため心筋細胞の表面は波打った形状をしている[17]。
細胞骨格は細胞内の足場であり、2つの主要な目的がある。1つ目は細胞内の構成要素の配置を安定させること、2つ目は細胞のサイズや形状を制御することである。1つ目の機能は生化学的過程に重要である。2つ目の機能は細胞の表面と体積の比率の決定に重要であり、興奮細胞の電気的特性に大きく影響する。さらに、細胞の標準的なサイズや形状からの逸脱は将来的に悪影響を及ぼす可能性がある[17]。
平滑筋細胞
平滑筋は筋原線維やサルコメアを持たず、そのため横紋筋ではない。平滑筋は胃、腸、膀胱、子宮など中空の器官の壁面や血管の壁面、呼吸器、泌尿器、生殖器の管に存在する。目では、毛様体筋が虹彩を広げたり縮めたりし、水晶体の形状を変化させている。皮膚では、立毛筋などの平滑筋細胞が寒さや恐怖に応答して毛を直立させる[18]。
平滑筋細胞は紡錘形で、中央部は幅広く、末端は細くなっている。単一の核を持ち、長さは30–200 μmである。これは骨格筋線維の数千分の一の長さである。細胞の直径も非常に小さいため、横紋筋細胞にあるT管は必要ない。平滑筋細胞にはサルコメアや筋原線維はないが、収縮タンパク質であるアクチンとミオシンは大量に含まれている。アクチンフィラメントは暗調小体(dense body、サルコメアのZ線に似ている)によって筋鞘に固定されている[18]。
発生
筋芽細胞はさまざまな筋細胞種へと分化する胚性前駆細胞である[19]。分化は、MyoD、Myf5、ミオゲニン、MRF4などの筋原性調節因子によって調節される[20]。GATA4とGATA6も筋細胞の分化に関与している[21]。
骨格筋線維は筋芽細胞の融合によって形成される。そのため、筋線維は筋核と呼ばれる複数の核を持つ細胞から構成される。筋核は各筋芽細胞の細胞核に由来するものである。筋芽細胞の融合は骨格筋特異的であり、心筋や平滑筋ではみられない。
骨格筋中で筋線維を形成しない筋芽細胞は衛星細胞へと脱分化する。こうした衛星細胞は骨格筋線維に近接したまま、筋鞘と筋内膜(筋束を個々の筋線維へ分離している結合組織)の基底膜の間に位置する[22]。筋形成を再活性化するためには、衛星細胞を刺激して新たな線維へと分化させる必要がある。
筋芽細胞や衛星細胞などその派生細胞は、多能性幹細胞の分化誘導によりin vitroで作り出すことができる[23]。
キンドリン-2は筋形成時の筋細胞の伸長に関与している[24]。
機能
横紋筋の収縮
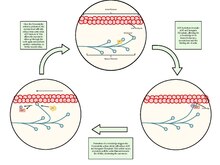
骨格筋の収縮
筋収縮時には、太いフィラメントと細いフィラメントはATPを利用して互いに滑り合い、これによってZ線は互いに引き寄せられる。この機構は滑り説と呼ばれている。全てのサルコメアが収縮することで、筋線維全体が収縮する。この筋細胞の収縮は細胞膜上の活動電位によって開始される。活動電位の筋細胞の表面から内部への伝導には、細胞膜と連続しているT管が利用される。筋小胞体は嚢状の膜であり、T管と接触しているが別個のものである。筋小胞体は各サルコメアを巻き、Ca2+で満たされている[25]。
筋細胞の興奮はそのシナプスである神経筋接合部での筋細胞の脱分極によって引き起こされ、活動電位が発生する。各筋線維には神経筋接合部が1つ存在し、1つの遠心性神経からのみの入力を受ける。遠心性神経の活動電位により、神経伝達物質であるアセチルコリンが放出される[26]。
放出されたアセチルコリンはシナプスを拡散し、筋鞘上の受容体に結合する。これによって筋鞘上を伝導するインパルスが開始される[27]。
活動電位が筋小胞体に到達すると、カルシウムチャネルからCa2+の放出が開始される。Ca2+は筋小胞体からサルコメアの双方のフィラメントへ流れる。これによってフィラメントはスライディングを開始し、サルコメアは短くなる。この過程には大量のATPが必要であり、全てのミオシン頭部の結合と解離の双方に利用される。Ca2+は筋小胞体へ非常に迅速に能動輸送によって送り返され、太いフィラメント細いフィラメントとの相互作用が遮断される。これによって筋細胞は弛緩する[27]。
筋収縮の様式には、いくつかのタイプが存在する。単収縮(攣縮、twitch)は、1回の刺激によって1回の収縮が行われる過程である。treppe/summationでは、筋肉は収縮を最大効率で開始するのではなく、反復刺激によって収縮強度を達成する過程であり、階段現象とも呼ばれる。強縮(tetanus)は、一連の迅速な刺激による筋肉の持続的な収縮であり、筋疲労を起こすまで継続される。等尺性収縮(isometric contraction)は筋肉の移動を伴わない骨格筋の収縮を指し、等張性収縮( isotonic contraction)は移動を伴う骨格筋の収縮を指す[27]。
心筋の収縮
洞房結節に位置する特殊な心筋細胞は、心拍数を制御する電気インパルスを発生させる。この電気インパルスは、心臓の刺激伝導系を介して残りの心筋全体の収縮を調整する。洞房結節の活動は、交感神経系と副交感神経系の双方の神経線維によって調節される。これらの神経系の神経線維は、それぞれ洞房結節での電気インパルスの発生率を高めたり低めたりする作用を果たす。
出典
- ^ Myocytes - MeSH・アメリカ国立医学図書館・生命科学用語シソーラス(英語)
- ^ a b Saladin, Kenneth S. (2011). Human anatomy (3rd ed.). New York: McGraw-Hill. pp. 72–73. ISBN 9780071222075
- ^ Scott, W.; Stevens, J.; Binder-Macleod, S. A. (2001-11). “Human skeletal muscle fiber type classifications”. Physical Therapy 81 (11): 1810–1816. ISSN 0031-9023. PMID 11694174.
- ^ “Does anyone know why skeletal muscle fibers have peripheral nuclei, but the cardiomyocytes not? What are the functional advantages?”. 2017年9月19日時点のオリジナルよりアーカイブ。2021年11月6日閲覧。
- ^ Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H.; Korol, Oksana; Johnson, Jody E.; Womble, Mark; Desaix, Peter (6 March 2013). "Cardiac muscle tissue" (Document).
{{cite document}}: Cite documentでは|publisher=が必須です。 (説明); 不明な引数|access-date=は無視されます。 (説明); 不明な引数|url=は無視されます。 (説明) - ^ “Muscle tissues”. 2015年10月13日時点のオリジナルよりアーカイブ。2015年9月29日閲覧。
- ^ “Atrial structure, fibres and conduction”. 2021年6月5日閲覧。
- ^ Saladin, Kenneth S. (2011). Human anatomy (3rd ed.). New York: McGraw-Hill. pp. 244–246. ISBN 9780071222075
- ^ “Structure of Skeletal Muscle | SEER Training”. training.seer.cancer.gov. 2021年11月6日閲覧。
- ^ Klein, CS; Marsh, GD; Petrella, RJ; Rice, CL (July 2003). “Muscle fiber number in the biceps brachii muscle of young and old men.”. Muscle & Nerve 28 (1): 62–8. doi:10.1002/mus.10386. PMID 12811774.
- ^ Cho, CH; Lee, KJ; Lee, EH (August 2018). “With the greatest care, stromal interaction molecule (STIM) proteins verify what skeletal muscle is doing.”. BMB Reports 51 (8): 378–387. doi:10.5483/bmbrep.2018.51.8.128. PMC 6130827. PMID 29898810.
- ^ Prasad, V; Millay, DP (2021-05-08). “Skeletal muscle fibers count on nuclear numbers for growth.”. Seminars in Cell & Developmental Biology. doi:10.1016/j.semcdb.2021.04.015. PMID 33972174.
- ^ a b c d Saladin, K (2012). Anatomy & Physiology: The Unity of Form and Function (6th ed.). New York: McGraw-Hill. pp. 403–405. ISBN 978-0-07-337825-1
- ^ Sugi, Haruo; Abe, T; Kobayashi, T; Chaen, S; Ohnuki, Y; Saeki, Y; Sugiura, S; Guerrero-Hernandez, Agustin (2013). “Enhancement of force generated by individual myosin heads in skinned rabbit psoas muscle fibers at low ionic strength”. PLOS ONE 8 (5): e63658. Bibcode: 2013PLoSO...863658S. doi:10.1371/journal.pone.0063658. PMC 3655179. PMID 23691080.
- ^ Charvet, B; Ruggiero, F; Le Guellec, D (April 2012). “The development of the myotendinous junction. A review.”. Muscles, Ligaments and Tendons Journal 2 (2): 53–63. PMC 3666507. PMID 23738275.
- ^ Bentzinger, CF; Wang, YX; Rudnicki, MA (1 February 2012). “Building muscle: molecular regulation of myogenesis.”. Cold Spring Harbor Perspectives in Biology 4 (2): a008342. doi:10.1101/cshperspect.a008342. PMC 3281568. PMID 22300977.
- ^ a b c “Healthy versus sick myocytes: metabolism, structure and function”. oxfordjournals.org/en. Oxford University Press. 2015年2月19日時点のオリジナルよりアーカイブ。2015年2月12日閲覧。
- ^ a b Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H.; Korol, Oksana; Johnson, Jody E.; Womble, Mark; Desaix, Peter (6 March 2013). "Smooth muscle" (Document).
{{cite document}}: Cite documentでは|publisher=が必須です。 (説明); 不明な引数|access-date=は無視されます。 (説明); 不明な引数|url=は無視されます。 (説明) - ^ Campbell, Neil A. (1999). Biology. Jane B. Reece, Lawrence G. Mitchell (5th ed ed.). Menlo Park, Calif.: Benjamin Cummings. p. 395. ISBN 0-8053-3044-5. OCLC 39981421
- ^ “Molecular mechanisms regulating myogenic determination and differentiation”. Front Biosci 5: D750–67. (2000). doi:10.2741/Perry. PMID 10966875.
- ^ “Loss of both GATA4 and GATA6 blocks cardiac myocyte differentiation and results in acardia in mice”. Dev. Biol. 317 (2): 614–9. (May 2008). doi:10.1016/j.ydbio.2008.03.013. PMC 2423416. PMID 18400219.
- ^ Zammit, PS; Partridge, TA; Yablonka-Reuveni, Z (November 2006). “The skeletal muscle satellite cell: the stem cell that came in from the cold.”. Journal of Histochemistry and Cytochemistry 54 (11): 1177–91. doi:10.1369/jhc.6r6995.2006. PMID 16899758.
- ^ “Differentiation of pluripotent stem cells to muscle fiber to model Duchenne muscular dystrophy”. Nat. Biotechnol. 33 (9): 962–9. (August 2015). doi:10.1038/nbt.3297. PMID 26237517.

- ^ “Kindlin-2 is required for myocyte elongation and is essential for myogenesis”. BMC Cell Biol. 9: 36. (2008). doi:10.1186/1471-2121-9-36. PMC 2478659. PMID 18611274.
- ^ “Structure and Function of Skeletal Muscles”. courses.washington.edu. 2015年2月15日時点のオリジナルよりアーカイブ。2015年2月13日閲覧。
- ^ “Muscle Fiber Excitation”. courses.washington.edu. University of Washington. 2015年2月27日時点のオリジナルよりアーカイブ。2015年2月11日閲覧。
- ^ a b c “Muscle Cell Anatomy & Function”. www.austincc.edu. 2015年9月23日時点のオリジナルよりアーカイブ。2015年2月12日閲覧。
外部リンク
 ウィキメディア・コモンズには、筋細胞に関するカテゴリがあります。
ウィキメディア・コモンズには、筋細胞に関するカテゴリがあります。- Structure of a Muscle Cell