
グリセロール3-リン酸
| L-グリセロール3-リン酸 (D-グリセロール1-リン酸) | |
|---|---|

| |
(R)-2,3-dihydroxypropyl dihydrogen phosphate | |
別称 1,2,3-propanetriol, 1-(dihydrogen phosphate), (2R)- D-glycerol 1-phosphate L-glycerol 3-phosphate L-α-glycerophosphate L-α-phosphoglycerol | |
| 識別情報 | |
| CAS登録番号 | 17989-41-2 |
| PubChem | 439162 |
| 日化辞番号 | J4.456C |
| KEGG | C00093 |
| MeSH | Alpha-glycerophosphoric+acid |
| |
| 特性 | |
| 化学式 | C3H9O6P |
| モル質量 | 172.074 |
| 関連する物質 | |
| 関連物質 | グリセロール1-リン酸 (鏡像異性体) |
| 特記なき場合、データは常温 (25 °C)・常圧 (100 kPa) におけるものである。 | |
sn-グリセロール3-リン酸(グリセロール3-りんさん、sn-glycerol 3-phosphate, G3P)[1]は、リン酸とグリセロールから誘導されたリン酸エステルで、グリセロリン脂質の構成要素の一つである。同等に適切な名前として、グリセロ-3-リン酸(glycero-3-phosphate)、3-O-ホスホノグリセロール(3-O-phosphonoglycerol)、3-ホスホグリセロール(3-phosphoglycerol)がある[2]。また歴史的経緯によりL-グリセロール3-リン酸(L-glycerol 3-phosphate)、D-グリセロール1-リン酸(D-glycerol 1-phosphate)、L-α-グリセロリン酸(L-α-glycerophosphoric acid)とも呼ばれる[2]。
生合成と代謝
[編集]グリセロール3-リン酸は解糖系の中間代謝物のうち、ジヒドロキシアセトンリン酸(DHAP)がグリセロール-3-リン酸デヒドロゲナーゼによって還元されて生じる。ブドウ糖ではなく、アミノ酸やTCA回路中間体などから糖新生経路を通ってDHAPを作り、同様にグリセロール3-リン酸を得ることもできる。
 + NADH + H+ →
+ NADH + H+ → 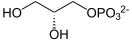 + NAD+
+ NAD+
脂肪を分解する場合には、生じたグリセロールをグリセロールキナーゼによってリン酸化してグリセロール3-リン酸とし、逆に解糖系や糖新生経路に供給することもできる。
グリセロール3-リン酸はグリセロリン脂質新規合成の出発物質の一つである。真核生物では小胞体膜またはミトコンドリア外膜に局在するグリセロール-3-リン酸アシル基転移酵素によって、グリセロール3-リン酸のsn-1位にアシル基がつきリゾホスファチジン酸となる。その後は小胞体でホスファチジン酸、1,2-ジアシルグリセロールと合成が進む。
- + Acyl-CoA → Lysophosphatidic acid + CoA
真菌ではグリセロール3-リン酸を脱リン酸化してグリセロールを生じるグリセロール脱リン酸化酵素があり、ブドウ糖から解糖系を経由してグリセロールを最終代謝物とするグリセロール発酵を行う場合がある。
- + H2O →
 + Pi
+ Pi
シャトル系
[編集]グリセロール-3-リン酸デヒドロゲナーゼは、ミトコンドリア内膜の膜間スペース側と細胞質の2ヶ所に存在している。またグリセロール3-リン酸(G3P)やジヒドロキシアセトンリン酸(DHAP)は分子量が小さく、ポリンを通ってミトコンドリア外膜を自由に通過できる。そこで、細胞質ではNADHを使ってDHAPをG3Pに還元し、ミトコンドリア内膜では電子伝達系のキノンを使ってG3PをDHAPに酸化するシャトル系が成立する。結果として、解糖系などにより細胞質側で生じたNADHをNAD+に再酸化し、またその還元当量を使ってプロトン勾配を形成してATP産生などに役立てることができる。こうしたシャトル系は他にも何種類かある。
鏡像異性体
[編集]グリセロール3-リン酸の鏡像異性体としてグリセロール1-リン酸がある。この2つは生体内で区別されており、一般的に活性を持つのはグリセロール3-リン酸であるが、グリセロール1-リン酸もアーキアの持つ特徴的なエーテル型脂質の骨格を与える代謝中間体として重要である。
脚注
[編集]この項目は、生化学に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:化学、プロジェクト:生命科学/Portal:化学、Portal:生物学)。 |