
ドクドニア・ドンヘンシス
| ドクドニア・ドンヘンシス | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 学名 | |||||||||||||||||||||
| Dokdonia donghaensis Yoon et al. 2005 |
ドクドニア・ドンヘンシス(Dokdonia donghaensis)は、海洋環境を好む絶対好気性でグラム陰性の光栄養細菌の一種である(→形態とゲノム)。海水培地中で生育可能な温度幅は広い(→生育環境)。バイオフィルムを形成する能力を持つ。当該バイオフィルムは、テトラサイクリンなどの抗生物質に対する本バクテリアの耐性を向上させる(→代謝)。
発見と分離[編集]
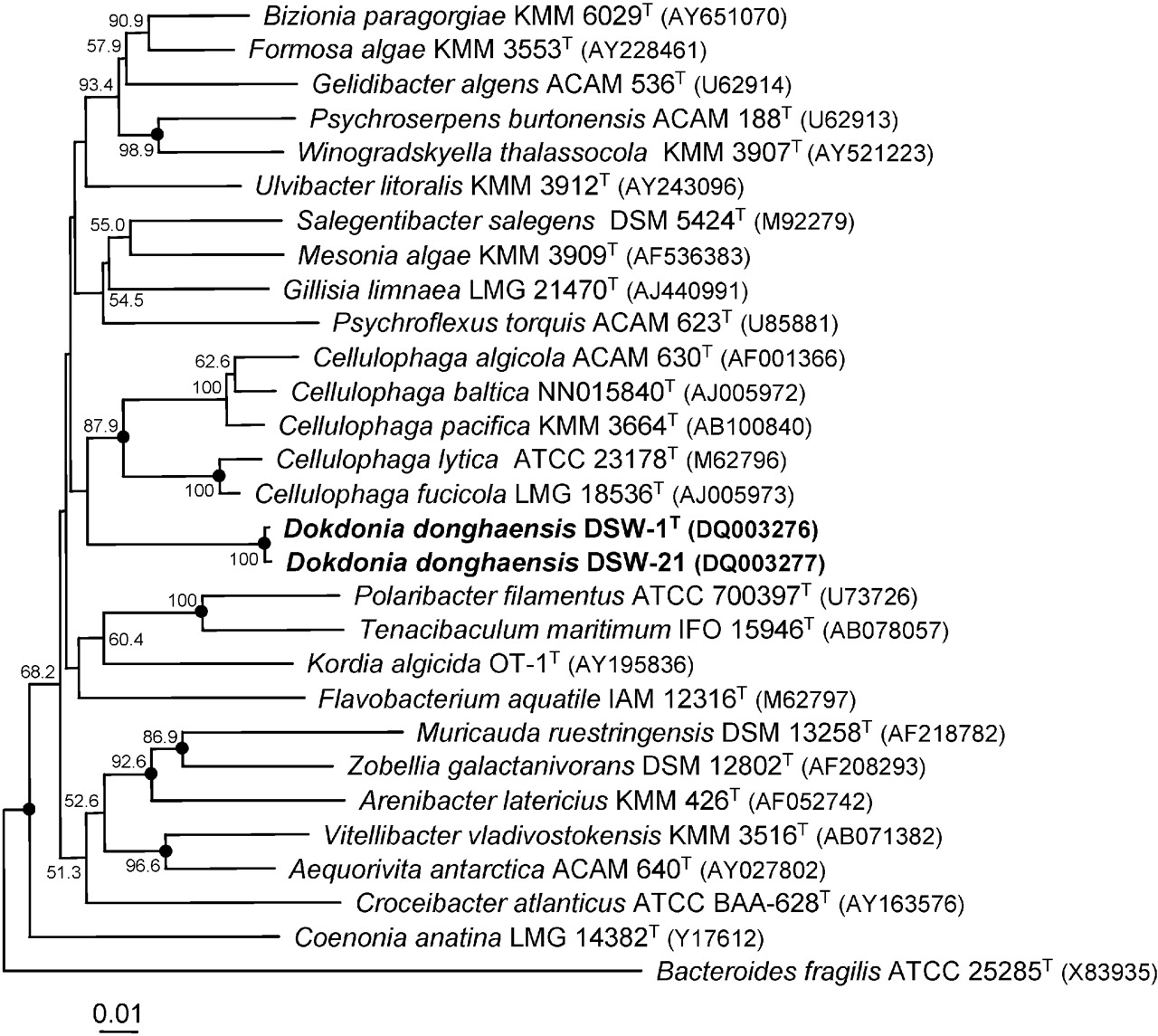
Yoon et al. (2005) は「東海にある韓国領の、独島と呼ばれる島から」、Dokdonia donghaensisを発見し、分離したと報告した[1]。分離の日付は特定されていない。本細菌の16SリボソームRNAの塩基配列に基づく系統解析によると、DSW-1T と DSW-21 の2系統と近い関係にあることがわかった。この2つの系統は、Cytophaga類縁細菌群(Cytophaga-Flavobacterium-Bacteroides (CFB) group)に分類され、海洋環境において優勢な属である[2][3]。セルゲイ・ヴィノグラツキイは、Cytophaga類縁細菌群が、形態こそ多様であるものの、単細胞生物であり、滑走を行い、芽胞を作らず、グラム陰性桿菌であることが共通していることを細菌研究史における早期に記述している。多くのCytophaga類縁細菌群の菌叢は、同細菌群とフラヴォバクテリアにしか見られないフレキシルビン型色素に由来する生物学的色素を持つ。Cytophaga類縁細菌群は化学合成有機栄養生物であり、キチン質やセルロースのような生体高分子(バイオポリマー)を分解できる[4]。
生息環境[編集]
D. donghaensis は好塩性であり海洋環境に生息する。生息可能温度は 4 °C から 35 °C まで。生長に適した温度は 30 °C。生長に適した pH は 7-8 であるが 5.5 より低い pH でも生長が観察された[5]。
形態とゲノム[編集]
Kim et al. (2016) による「東海(韓国海)中にある独島沿岸の海水から分離された、プロテオロドプシンを含む海洋性のフラヴォバクテリウム・ドクドニア・ドンヘンシス DSW-1T 株の完全なゲノム配列」と題した論文によると、D. donghaensis は、桿状の外観を呈し、非運動性で、芽胞を作らないグラム陰性菌の一種である[6]。海水寒天培地上で培養させることが可能であり、その菌叢は直径 1-2 mm の円形を呈する[6]。また、わずかに盛り上がり、ツルツルしており、光を反射して黄色に輝く[6]。D. donghaensisの DSW-1T 菌株は、その遺伝情報全体(ゲノム)の中に約3,923,666の塩基対を有する[6]。D. donghaensis のデオキシリボ核酸のグアニン+シトシン含量は、38% である[6]。
Microbiology Society が系統樹を提供している。IJSEM への登録は、「独島東海」と名づけられた本種のほか、「独島韓国(Dokdonella koreensis)」、「ヴァージバシラス独島(Virgibacillus dokdonensis)」、「マリバクター独島(Maribacter dokdonensis)」、「マリノモナス独島(Marinomonas dokdonensis)」と同時に行われた[7]。このような韓国籍人による新種の微生物の命名の動向については、何らかの地政学的目的があると推定されている[8]。
{kind=link}
代謝[編集]
D. donghaensis は光によって駆動されるナトリウムイオン輸送を行う。この光駆動性プロトンポンプは、プロテオロドプシンと呼ばれている。プロテオロドプシンは光を、微生物の生長に必要とするエネルギーへと変換できる。そのため、微生物の生存率を増加させる[9]。D. donghaensis は光を栄養にするので、光栄養生物である。D. donghaensis は、カタラーゼやオキシダーゼといった酵素を分泌することができる[10]。D. donghaensis は偏性好気菌(strict aerobe)であるため、酸素を最終電子受容体として用いる。D. donghaensis は、ペプトンとトリプトン培地を唯一の炭素・窒素源として生育可能である。D. donghaensis は、塩化ナトリウムがまったくないか、あるいは溶液濃度で 7% (w/v)を越えた環境下では生育不能である。生長には 2% (w/v) NaCl であることが好ましい。D. donghaensis は、抗生物質であるテトラサイクリン(テトラサイクリン系抗生物質)とカルベニシリンに弱いが、抗生物質が透過しにくい粘性のあるバイオフィルムを形成する能力を有する[11]。
バイオフィルムの形成[編集]
D. donghaensis は、海洋環境においてバイオフィルムを形成することができる。これは環境負荷から保護された状態で生長を図るという生存戦略の一つである。海洋細菌がバイオフィルムを形成する目的は、抗菌因子、乾燥、接触による細胞の損傷に対する抵抗力を増加させることにある[12][13][14][15]。バイオフィルムにより、微生物は細胞外高分子物質(EPS: Extracellular polymeric substances)を排泄することで海面に居続けることができる。海面に固着する海洋細菌は、宿主特異的で空間的に構築された、安定性の良好な微生物群落を形成する[16][17][18]。
病理学的知見[編集]
2017年1月時点において、D. donghaensis がヒト病原体であるか否かは確認されていない[要出典]。
出典[編集]
- ^ Yoon, JH; Kang, SJ; Lee, CH; Oh, TK (2005). “Dokdonia donghaensis gen. nov., sp. nov., isolated from sea water”. Int J Syst Evol Micr 55 (6).
- ^ Bowman, JP; McCammon, SA; Brown, MV; Nichols, DS; McMeekin, TA (1997). “Diversity and association of psychrophilic bacteria in Antarctic sea ice”. Appl Environ Microbiol 63.
- ^ Glöckner, FO; Fuchs, BM; Amann, R (1999). “Bacterioplankton compositions of lakes and oceans: a first comparison based on fluorescence in situ hybridization”. Appl Environ Microbiol 65.
- ^ Kirchman, DL (2002). “The ecology of Cytophaga-Flavobacteria in aquatic environments”. FEMS Microbiol Ecol 39.
- ^ Yoon, JH; Kang, KH; Park, YH (2003). “Psychrobacter jeotgali sp. nov., isolated from jeotgal, a traditional Korean fermented seafood”. Int J Syst Evol Micr 53 (2).
- ^ a b c d e Kim, K; Kwon, SK; Yoon, JH; Kim, JF (2016). “Complete genome sequence of the proteorhodopsin-containing marine flavobacterium Dokdonia donghaensis DSW-1T, isolated from seawater off Dokdo in the East Sea (Sea of Korea)”. Genome Announc 4 (4). doi:10.1128/genomeA.00804-16. PMID 27491981.
- ^ “「微生物の名前で‘独島は韓国領’知らせる」”. 中央日報日本語版. 中央日報 (2005年6月9日). 2017年7月3日閲覧。
- ^ Fred A. Rainey; Aharon Oren, eds (2011). Taxonomy of Prokaryotes. Academic Press. pp. 473 2017年7月3日閲覧。 p453.
- ^ DeLong, EF; Beja, O (2010). “The light-driven proton pump proteorhodopsin enhances bacterial survival during tough times”. PLoS Biol 8 (4).
- ^ Wang, Y; Zhou, C; Ming, H; Kang, J; Chen, H; Jing, C; Feng, H; Chang, Y et al. (2016). “Pseudofulvibacter marinus sp. nov., isolated from seawater”. Int J Syst Evol Micr 66 (3).
- ^ Burmølle, M; Webb, JS; Rao, D; Hansen, LH; Sørensen, SJ; Kjelleberg, S (2006). “Enhanced biofilm formation and increased resistance to antimicrobial agents and bacterial invasion are caused by synergistic interactions in multispecies biofilms”. Appl Environ Microbiol 72 (6). doi:10.1128/AEM.03022-05. PMID 16751497.
- ^ Fux, CA; Costerton, JW; Stewart, PS; Stoodley, P (2005). “Survival strategies of infectious biofilms”. Trends Microbiol 13 (1).
- ^ Mah, TF; O'Toole, GA (2001). “Mechanisms of biofilm resistance to antimicrobial agents”. Trends Microbiol 9.
- ^ Jefferson, KK (2004). “What drives bacteria to produce a biofilm?”. FEMS Microbiol Lett 236.
- ^ Matz, C; Kjelleberg, S (2005). “Off the hook—how bacteria survive protozoan grazing”. Trends Microbiol 13.
- ^ Pasmore, M; Costerton, JW (2003). “Biofilms, bacterial signaling, and their ties to marine biology”. J Ind Microbiol Biotechnol 30.
- ^ Taylor, MW; Schupp, PJ; Dahllöf, I; Kjelleberg, S; Steinberg, PD (2004). “Host specificity in marine sponge-associated bacteria, and potential implications for marine microbial diversity”. Environ Microbiol 6.
- ^ Bhadury, P; Wright, PC (2004). “Exploitation of marine algae: biogenic compounds for potential antifouling applications”. Planta 219.